
















Goldcrest Regulus regulus Scientific name definitions
Text last updated January 16, 2013
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Albanian | Mbretëthi |
| Arabic | صعو أصفر العرف |
| Armenian | Դեղնագլուխ արքայիկ |
| Asturian | Reyñn nortiegu |
| Azerbaijani | Sarıbaş kralcıq |
| Basque | Mendi-erregetxoa |
| Bulgarian | Жълтоглаво кралче |
| Catalan | reietó eurasiàtic |
| Chinese | 戴菊鳥 |
| Chinese (Hong Kong SAR China) | 戴菊 |
| Chinese (SIM) | 戴菊 |
| Croatian | zlatoglavi kraljić |
| Czech | králíček obecný |
| Danish | Fuglekonge |
| Dutch | Goudhaan |
| English | Goldcrest |
| English (United States) | Goldcrest |
| Faroese | Gulkrýndur títlingskongur |
| Finnish | hippiäinen |
| French | Roitelet huppé |
| French (France) | Roitelet huppé |
| Galician | Estreliña dos piñeiros |
| German | Wintergoldhähnchen |
| Greek | Χρυσοβασιλίσκος |
| Hebrew | מלכילון האורנים |
| Hungarian | Sárgafejű királyka |
| Icelandic | Glókollur |
| Italian | Regolo |
| Japanese | キクイタダキ |
| Korean | 상모솔새 |
| Latvian | Zeltgalvītis |
| Lithuanian | Paprastasis nykštukas |
| Mongolian | Шар мэлзэн задуулай |
| Norwegian | fuglekonge |
| Persian | سسک تاج طلایی |
| Polish | mysikrólik |
| Portuguese (Portugal) | Estrelinha-de-poupa |
| Romanian | Aușel cu cap galben |
| Russian | Желтоголовый королёк |
| Serbian | Kraljić |
| Slovak | králik zlatohlavý |
| Slovenian | Rumenoglavi kraljiček |
| Spanish | Reyezuelo Sencillo |
| Spanish (Spain) | Reyezuelo sencillo |
| Swedish | kungsfågel |
| Turkish | Çalıkuşu |
| Ukrainian | Золотомушка жовточуба |
Regulus regulus (Linnaeus, 1758)
Definitions
- REGULUS
- regulus
The Key to Scientific Names
Legend Overview
Originally Appeared in

Field Identification
8·5–9·5 cm; 4·6–7·1 g. Tiny olive passerine , fairly compact in appearance, with distinctive face pattern. Male nominate race has bright lemon-yellow central crownstripe with rear feathers intense orange (orange visible only when crest raised), bordered by black lateral stripes, broad but diffuse white eyering (emphasizing dark eye), short black moustachial stripe; upperparts pale olive-green, two white wingbars, basal black patch on secondaries forming sharply contrasting rear bar, flight-feathers narrowly edged yellowish, tertials edged and tipped white; throat and underparts greyish-white, tinged yellowish (when plumage fresh); iris dark brown; bill thin and needle-like, black-brown; legs brown. Easily distinguished from R. ignicapilla by duller and less colourful plumage, black eyestripe lacking, black rear bar on wing broader and more sharply contrasted. Female has crown paler and lacking orange feathers. Juvenile is duller than adult, lacks bright crown feathers, has white eyering less conspicuous and with narrow dark brown interior ring. Races vary mainly in plumage coloration and size: coatsi has paler upperparts and broader white tips on upperwing-coverts than nominate, wing on average longer; inermis has more mottled, darker green-olive upperparts and more brownish-olive underparts than nominate, longer bill; azoricus is similar to previous, but more olive-buff to yellowish below; sanctaemariae is much paler than previous two, more pale yellowish-olive above, brighter creamy white below; <em>teneriffae</em> is slightly smaller than nominate but longer-billed , generally brighter, has black lateral crownstripes meeting on forehead, stronger orange on crown of male; ellenthalerae differs from last in having finer bill, underparts less brownish, male with central crownstripe paler; buturlini is duller and greyer above and less yellowish below than nominate, wingbars more conspicuous; hyrcanus is darker than previous, greyer than nominate; tristis is distinctive, much duller and greyer above, with lower back and rump greyish-olive, black lateral crownstripes greatly reduced and browner or even lacking, male crown yellow or orange-yellow (never orange), female crown patch smaller or absent; himalayensis is rather dark olive-greyish; sikkimensis is darker and greener above than previous; yunnanensis is darkest race, nape slate-grey; <em>japonensis</em> has nape and head side pure grey, darker and greener above, white wing-covert tips somewhat broader, brownish tinge below (in fresh plumage more ochre).
Systematics History
Editor's Note: This article requires further editing work to merge existing content into the appropriate Subspecies sections. Please bear with us while this update takes place.
Canary Is race teneriffae (with, by implication, recently described race ellenthalerae) treated variously as a race of R. ignicapilla or as a separate species, but acoustic and molecular markers indicate that Canarian populations belong with W Palearctic complex of races of present species that also includes nominate and the three Azores races (inermis, azoricus, sanctaemarie). These W races are opposed to a complex of E Asian races, and acoustic and genetic divergence between the two groups is evidently high. Further, on Azores, race azoricus (from São Miguel) apparently does not represent a monophyletic taxon but is evidently separated into an E group, with affiliation to sanctaemariae (from Santa Maria), and a W population that genetically belongs to inermis (W islands of Azores). In addition, geographical variation partly clinal, birds becoming darker and slightly larger from W to E in Palearctic; buturlini and hyrcanus possibly not truly separable. Another morphological cline in S Asia, from dark greyish plumage in Himalayan himalayensis towards brighter colours in Chinese sikkimensis and yunnanensis; these three taxa not vocally or genetically differentiated, either, and subspecific separation possibly unreliable. Taxonomy of this species clearly in need of revision, but must be based on thorough revision of genus. Otherwise, proposed races anglorum (from Britain) and interni (from Corsica) synonymized with nominate, although latter, at least, exhibits slight acoustic differentiation from neighbouring continental populations. Fourteen subspecies recognized.Subspecies
Goldcrest (European) Regulus regulus [regulus Group]


Regulus regulus regulus (Linnaeus, 1758)
Definitions
- REGULUS
- regulus
The Key to Scientific Names
Legend Overview
Distribution
Regulus regulus coatsi Sushkin, 1904
Definitions
- REGULUS
- regulus
- coatesi / coatsi
The Key to Scientific Names
Legend Overview
Distribution
Regulus regulus buturlini Loudon, 1911
Definitions
- REGULUS
- regulus
- buturlini
The Key to Scientific Names
Legend Overview
Regulus regulus hyrcanus Zarudny, 1910
Definitions
- REGULUS
- regulus
- hyrcana / hyrcanus
The Key to Scientific Names
Legend Overview
Regulus regulus tristis Pleske, 1892
Definitions
- REGULUS
- regulus
- tristis
The Key to Scientific Names
Legend Overview
Goldcrest (western Canary Islands) Regulus regulus ellenthalerae Scientific name definitions
Distribution
Regulus regulus ellenthalerae Päckert et al., 2006
Definitions
- REGULUS
- regulus
- ellenthalerae
The Key to Scientific Names
Legend Overview
Goldcrest (Tenerife) Regulus regulus teneriffae Scientific name definitions

Distribution
Regulus regulus teneriffae Seebohm, 1883
Definitions
- REGULUS
- regulus
- tenerifae / teneriffae
The Key to Scientific Names
Legend Overview
Goldcrest (Sao Miguel) Regulus regulus azoricus Scientific name definitions

Distribution
Regulus regulus azoricus Seebohm, 1883
Definitions
- REGULUS
- regulus
- azorensis / azorica / azoricus
The Key to Scientific Names
Legend Overview
Goldcrest (Santa Maria) Regulus regulus sanctaemariae Scientific name definitions
Distribution
Regulus regulus sanctaemariae Vaurie, 1954
Definitions
- REGULUS
- regulus
- sanctaemariae
The Key to Scientific Names
Legend Overview
Goldcrest (Western Azores) Regulus regulus inermis Scientific name definitions
Distribution
Regulus regulus inermis Murphy & Chapin, 1929
Definitions
- REGULUS
- regulus
- inerme / inermis
The Key to Scientific Names
Legend Overview
Goldcrest (Asian) Regulus regulus [himalayensis Group]


Distribution
Regulus regulus himalayensis Bonaparte, 1856
Definitions
- REGULUS
- regulus
- himalayensis
The Key to Scientific Names
Legend Overview
Distribution
Regulus regulus sikkimensis Meinertzhagen & Meinertzhagen, 1926
Definitions
- REGULUS
- regulus
- sikkimensis
The Key to Scientific Names
Legend Overview
Distribution
Regulus regulus japonensis Blakiston, 1862
Definitions
- REGULUS
- regulus
- japanensis / japanicus / japensis / japonensis / japonica / japonicum / japonicus
The Key to Scientific Names
Legend Overview
Distribution
S Shaanxi and Sichuan S to SE Tibet and N Yunnan (China) and NE Myanmar.
Regulus regulus yunnanensis Rippon, 1906
Definitions
- REGULUS
- regulus
- yunnanense / yunnanensis
The Key to Scientific Names
Legend Overview
Distribution
Editor's Note: Additional distribution information for this taxon can be found in the 'Subspecies' article above. In the future we will develop a range-wide distribution article.
Habitat
Boreal forests dominated by Norway spruce (Picea abies) and fir (Abies); on Atlantic islands in laurel (Laurus) forest or pine (Pinus) stands with well-developed undergrowth of tree-heath (Erica), but also in conifer stands of indigenous short-leaved juniper (Juniperus brevifolia) or introduced Japanese cedar (Cryptomeria japonica). Outside breeding season, and especially during migration, also invades mixed and deciduous forests, shrub vegetation and gardens and parks. Breeds at up to 2200 m in Swiss Alps (nominate race), to 2000 m in Caucasus (buturlini), to 2400 m in Ala Tau (tristis), to 2600 m in Japan Alps and on Mt Fuji (japonensis) and to 4000 m in Himalayas (himalayensis).
Movement
Chiefly migratory. Only N Fennoscandian breeding grounds (mainly in Finland) completely abandoned by entire population; in C Europe, local breeding populations depart from beginning of Aug and, by mid-Dec, have been replaced by wintering flocks from N Europe. Two main migration routes, one SW along coastlines of Norway, Denmark and Netherlands into W & S Europe, and another E route from Scandinavia and the Baltic states to Poland and S to Balkans; winters in large parts of S Europe, including Mediterranean islands (Corsica, Sardinia, Sicily, Malta, Cyprus), but only limited wintering in N Africa (regular in small numbers in Tunisia, 4 records from Morocco). C Asian populations (of race tristis) descend to foothills in winter, moving SW beyond breeding boundaries to Tadjikistan, Turkmenistan and Afghanistan, with occasional records as far W as Iran. E Asian japonensis winters in Korea, E China and Ryukyu Is, and occasionally reaches Taiwan. In Himalayas, makes post-breeding descent to lower levels; race sikkimensis from NC China partially moves to Yangtze Valley in winter. Resident populations on Azores, Canary Is and Corsica.
Diet and Foraging
Arthropods. Generally adapted to small-sized prey, such as springtails (Collembola), aphids (Aphidoidea), small moths (Lepidoptera) and tiny spiders (Araneae). Strong preference for springtails (Collembola), especially during non-breeding season, when bark-living species of genus Entomobrya important. In Canary Is less selective in terms of prey size, commonly feeds on long-legged arthropods such as orthopterans; in captivity readily took phasmids, which generally shunned by nominate race in mainland Europe. With decreasing food availability during migration, larger species, such as adult winter moths (Operophtera brumata), also taken. Largest items usually beaten repeatedly against a branch. Forages mostly by clinging and hanging from twigs. In coniferous habitats , prefers to forage in dense branches, moving up and down vertical twigs; exploits underside of branches more often and more intensely than does R. ignicapilla. Also hovers at outer tips of branches, picking prey from substrate or in flight. Sometimes hovers at spider webs, from which picks trapped small insects such as aphids and psyllids (Psyllidae); rarely, becomes entangled in web and may perish. Outside breeding season often in small, loose groups, frequently with mixed-species flocks.
Sounds and Vocal Behavior
Territorial song composed of a stereotype high-pitched main part and a shorter, highly variable terminal flourish, each male possessing repertoire of up to 28 different terminal motifs; geographical variation extremely high, with distinct regional dialects; main part of song either a rapid trill (“himalayensis group”, Hokkaido population of japonensis, azoricus, sanctaemariae, teneriffae) or a rhythmic alternation of high-frequency and low-frequency notes “tee-de-dee tee-de-dee” (nominate , continental japonensis, inermis, other song types of azoricus and sanctaemariae). Calls are high-pitched and quiet “seeeh” or “zick”, sometimes louder.
Breeding
Beginning of nesting activities from end of Mar, nest records mainly from first week Apr; two broods per breeding season. Nest built by both sexes, a typical regulid cup-nest of three layers, inner layer padded out with large number of small feathers (up to 2500 in single nest), strong coat of lichens (especially of genus Parmelia) in outer layer, suspended 4–14 m above ground in vertical twigs of conifer; on Atlantic islands, nest typically built in species of tree-heath, e.g. incorporated in horizontal twigs of Erica arborea in Canaries (race teneriffae and ellenthalerae), also a few nest recorded in laurel trees. Clutch 6–13 eggs , second clutches slightly smaller than first; incubation by female, period 14–17 days (in captivity, average 15 days); chicks fed by both sexes, nestling period 17–22 days (in captivity, 19–21 days after hatching); fledglings fed by parents for 12–18 days after having left nest.
Conservation Status
Not globally threatened (Least Concern). Common to locally abundant. Marked range extension after planting of spruce forests in C Europe in 19th century. Numbers fluctuate, depending largely on severity of winter weather; estimates of European population include up to several tens of millions of breeding pairs in “good” years. Some island races may require further consideration because of ongoing loss of suitable breeding habitats, such as the laurel forests of Canary Is (<em>teneriffae</em> and ellenthalerae) or those of the Azores; in the latter islands, race sanctaemariae has only a very restricted forest area on the C mountain (Pico Alto) of Santa Maria. Although harsh winter conditions, especially if they persist for lengthy periods over wide aeas of the non-breeding range, can seriously reduce population levels, this species is capable of recovering its numbers within only a few breeding seasons.
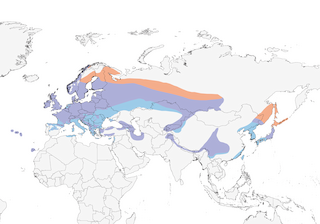
- Year-round
- Migration
- Breeding
- Non-Breeding


















{kind=link}