












Great Spotted Woodpecker Dendrocopos major Scientific name definitions
- LC Least Concern
- Names (45)
- Subspecies (14)
Hans Winkler, David Christie, and Guy M. Kirwan
Version: 1.0 — Published March 4, 2020
Text last updated September 6, 2015
Text last updated September 6, 2015
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Albanian | Qukapiku larosh i madh |
| Armenian | Խայտաբղետ փայտփոր |
| Asturian | Picatueru comñn |
| Azerbaijani | İri ağacdələn |
| Basque | Okil handia |
| Bulgarian | Голям пъстър кълвач |
| Catalan | picot garser gros |
| Chinese (SIM) | 大斑啄木鸟 |
| Croatian | veliki djetlić |
| Czech | strakapoud velký |
| Danish | Stor Flagspætte |
| Dutch | Grote Bonte Specht |
| English | Great Spotted Woodpecker |
| English (United States) | Great Spotted Woodpecker |
| Faroese | Stór flekkuspæta |
| Finnish | käpytikka |
| French | Pic épeiche |
| French (France) | Pic épeiche |
| Galician | Picapau común |
| German | Buntspecht |
| Greek | Πευκοδρυοκολάπτης |
| Hebrew | נקר עקוד |
| Hungarian | Nagy fakopáncs |
| Icelandic | Barrspæta |
| Italian | Picchio rosso maggiore |
| Japanese | アカゲラ |
| Korean | 오색딱다구리 |
| Latvian | Dižraibais dzenis |
| Lithuanian | Didysis margasis genys |
| Mongolian | Алаг тоншуул |
| Norwegian | flaggspett |
| Persian | دارکوب خالدار بزرگ |
| Polish | dzięcioł duży |
| Portuguese (Angola) | Great Spotted Woodpecker |
| Portuguese (Portugal) | Pica-pau-malhado |
| Romanian | Ciocănitoare pestriță mare |
| Russian | Большой пёстрый дятел |
| Serbian | Veliki detlić |
| Slovak | ďateľ veľký |
| Slovenian | Veliki detel |
| Spanish | Pico Picapinos |
| Spanish (Spain) | Pico picapinos |
| Swedish | större hackspett |
| Turkish | Orman Alaca Ağaçkakanı |
| Ukrainian | Дятел звичайний |
Dendrocopos major (Linnaeus, 1758)
PROTONYM:
Picus major
Linnaeus, 1758. Systema Naturæ per Regna Tria Naturæ, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis. Tomus I. Editio decima, reformata 1, p.114.
TYPE LOCALITY:
Europe == Sweden ex Fn. Suec.
SOURCE:
Avibase, 2023
Definitions
- DENDROCOPOS
- major
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
Originally Appeared in

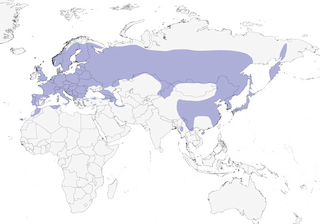
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Great Spotted Woodpecker















