














Greater Whitethroat Curruca communis Scientific name definitions
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Afrikaans | Witkeelsanger |
| Albanian | Bilbilthi i përhimë |
| Arabic | هازجة بيضاء الحنجرة الكبرى |
| Armenian | Մոխրագույն շահրիկ |
| Asturian | Papuda les sebes |
| Azerbaijani | Boz silvi |
| Basque | Sasi-txinboa |
| Bulgarian | Голямо белогушо коприварче |
| Catalan | tallareta comuna |
| Chinese (SIM) | 灰白喉林莺 |
| Croatian | grmuša pjenica |
| Czech | pěnice hnědokřídlá |
| Danish | Tornsanger |
| Dutch | Grasmus |
| English | Greater Whitethroat |
| English (India) | Greater Whitethroat (Common Whitethroat) |
| English (Kenya) | Common Whitethroat |
| English (UK) | Common Whitethroat |
| English (United States) | Greater Whitethroat |
| Faroese | Tyrniljómari |
| Finnish | pensaskerttu |
| French | Fauvette grisette |
| French (France) | Fauvette grisette |
| Galician | Papuxa común |
| German | Dorngrasmücke |
| Greek | Θαμνοτσιροβάκος |
| Hebrew | סבכי קוצים |
| Hungarian | Mezei poszáta |
| Icelandic | Þyrnisöngvari |
| Italian | Sterpazzola |
| Japanese | ノドジロムシクイ |
| Latvian | Brūnspārnu ķauķis |
| Lithuanian | Rudoji devynbalsė |
| Mongolian | Шөвгөр зэржгэнэ |
| Norwegian | tornsanger |
| Persian | سسک گلوسفید معمولی |
| Polish | cierniówka |
| Portuguese (Angola) | Papa-amoras |
| Portuguese (Portugal) | Papa-amoras-comum |
| Romanian | Silvie de câmp |
| Russian | Серая славка |
| Serbian | Obična grmuša |
| Slovak | penica obyčajná |
| Slovenian | Rjava penica |
| Spanish | Curruca Zarcera |
| Spanish (Spain) | Curruca zarcera |
| Swedish | törnsångare |
| Turkish | Ak Gerdanlı Ötleğen |
| Ukrainian | Кропив'янка сіра |
Curruca communis (Latham, 1787)
Definitions
- CURRUCA
- curruca
- communis
- Communis
The Key to Scientific Names
Legend Overview
Field Identification
14 cm; 14–17 g. A medium-sized Curruca, quite slim, with rather large head and long tail, and moderately long wings with noticeable rufous wingpanel. Male nominate race has ash-grey to pure grey head, subtle and ill-defined narrow greyish supercilium, indistinct white eyering ; upperparts brown to grey-brown, uppertail-coverts greyer, tertials, inner secondaries and larger upperwing-coverts with distinct black-brown centres and pale rufous fringes and tips (obvious rufous wingpanel); tail dark brown, outermost feathers with whitish edges and tips; chin and upper throat pure white; whitish below, lower throat and breast with variable suffusion of pinkish-buff, buffish on flanks; iris pale yellow to light brown; bill with dark grey culmen and tip, pinkish-horn basal area; legs pinkish-brown, often with yellow tinge. Female is similar to male but duller and browner overall, especially on crown and cheeks, has less strong rufous tones on upperwing-coverts, duller whitish on tail edges, lacks pinkish tinge below, has iris duller, brown to olive-grey (some females have grey crown and some pinkish below, almost identical to younger males). Juvenile is rather uniform buff-brown above, chin and throat off-white, underparts mostly dull buff, edges and tips of outermost rectrices sandy buff, iris dark greyish-olive; first-winter mostly like autumn female, flight-feathers and bare parts largely as juvenile. Race volgensis is similar to nominate but slightly larger, wing more pointed and longer, paler and greyer above, male whiter below and with pale pinkish suffusion, female duller (but probably mostly indistinguishable from nominate); <em>icterops</em> has wing rather pointed and relatively long, male crown purer grey and considerably darker than nominate, is darker and greyer above, mantle and scapulars greyish-brown, wingpanel duller and indistinct, underparts whiter and with lesser amount of pinkish (undertakes characteristic complete prenuptial moult in winter quarters); rubicola is similar to previous (wing also pointed and longer, has same complete winter moult), but with more grey above and paler overall, with underparts whiter.
Systematics History
Editor's Note: This article requires further editing work to merge existing content into the appropriate Subspecies sections. Please bear with us while this update takes place.
Geographical variation largely clinal, but races form two rather well-differentiated groups: NW group contains nominate race and volgensis, which intergrade widely in E Europe, and SE group comprises icterops and rubicola; groups intergrade in SE Europe, Turkey, Caucasus and Altai. Proposed races jordansi (from British Is except SE & E England), cinerea (Scandinavia, Germany S to N Greece) and hoyeri (E from E Poland) synonymized with nominate, and traudeli (E Asia Minor) included in icterops. Four subspecies recognized.Subspecies
Curruca communis communis Scientific name definitions


Distribution
Curruca communis communis (Latham, 1787)
Definitions
- CURRUCA
- curruca
- communis
- Communis
The Key to Scientific Names
Legend Overview
Curruca communis volgensis Scientific name definitions
Distribution
Curruca communis volgensis (Domaniewski, 1915)
Definitions
- CURRUCA
- curruca
- communis
- Communis
- volgae / volgensis
The Key to Scientific Names
Legend Overview
Curruca communis icterops Scientific name definitions
Distribution
Curruca communis icterops (Ménétries, 1832)
Definitions
- CURRUCA
- curruca
- communis
- Communis
- icterops
The Key to Scientific Names
Legend Overview
Curruca communis rubicola Scientific name definitions


Distribution
Curruca communis rubicola (Stresemann, 1928)
Definitions
- CURRUCA
- curruca
- communis
- Communis
- rubicola
- Rubicola
The Key to Scientific Names
Legend Overview
Distribution
Editor's Note: Additional distribution information for this taxon can be found in the 'Subspecies' article above. In the future we will develop a range-wide distribution article.
Habitat
Open country in wide variety of landscapes, mostly sunlit areas with scattered bushes and shrubs close to grassy patches; avoids densely vegetated areas, closed woodland and formations taller than 3–4 m. Breeds in plantation clearings, orchards, hedgerows along roads or railway lines, shrubs near watercourses, pastures with junipers (Juniperus); in steppes also in halophytic vegetation such as Halimodendron or Artemisia; also in hedges around field crops, especially those of appropriate height such as cereals or lucerne (Medicago). Favours lowlands, but breeds at 1500–2000 m in Caucasus, up to 1600 m in the Alps, and above 2000 m in Moroccan Atlas; recorded at c. 3200 m in Kazakhstan (race rubicola). On African non-breeding grounds, occupies grassy steppes with sparse acacias (Acacia) and other trees (Balanites, Ziziphus), semi-arid savanna, and evergreen and semi-evergreen trees with scrub; in Kenya sometimes shares acacias and Commiphora woodland above 700 m with C. nisoria, but selects comparatively less arid areas and with lower cover; in S Africa, overlaps in range with Sylvia borin and attains highest densities in Baikiaea woodland. During migration in E Africa, also visits denser bushes, hedges and highlands up to 2000 m; recorded in Zambia and Zimbabwe in dry acacia shrub and secondary vegetation with Lantana.
Movement
Migratory. All populations spend non-breeding season in sub-Saharan Africa , from Senegal E to Sudan and N Somalia and S to South Africa. In autumn, European continental populations exhibit a migratory divide around 10º E (C Germany), W populations migrating SW through Iberian Peninsula and wintering mostly in W Africa, whereas E ones fly through Italy and coast of E Mediterranean Sea and winter in C & W Africa. British breeders apparently first undertake SE-oriented flight to reach France and later migrate SW through Iberia, where can stop over for some days before crossing Sahara to W Africa. The two E races (icterops, rubicola) move through Middle East to reach Sudan, Eritrea and Ethiopia in Aug–Sept, remaining there for some months (many individuals moult); in Nov–Dec continue S and arrive in S Africa from mid-Nov to late Dec; exact wintering ranges of each E race largely unknown. Spring passage noted in S Africa from mid-Mar to mid-Apr, at which period especially intense in Zambia and Malawi; also, marked passage in Ethiopia (mainly from late Mar onwards) and Eritrea (early May), and conspicuous in Somalia during May; in Sudan numbers lower than in autumn, suggesting more E migration in spring, but in N Africa more conspicuous than in autumn; spring passage recorded mainly late Mar to mid-May in Morocco and Algeria. Arrival on breeding grounds mainly in Apr and May.
Diet and Foraging
Well known; detailed studies based on analyses of both stomach contents and collar-samples carried out mostly in Europe. Invertebrates taken throughout year, include large number of insect orders and also other arthropods; fruits increase in significance outside breeding season, being particularly important during autumn migration and sometimes also in winter. During breeding season, adults fed mostly on beetles (especially of families Curculionidae and Chrysomelidae), bugs (Hemiptera), lepidopterans and hymenopterans; importance of beetles demonstrated by some studies elsewhere in breeding range; hemipterans the commonest prey in analyses of samples from E Germany and Ukraine. Changes throughout breeding season reported in study in NE Germany, probably reflecting local availability of prey abundance; beetles predominant in May, hymenopterans more common in late Aug and early Sept, and steady increase in consumption of fruits once autumn underway. Nestlings fed with comparatively softer prey than taken by adults, including more spiders (Araneae) and caterpillars and fewer beetles; samples from Sweden and Kazakhstan revealed dominance of lepidopterans (mainly larvae) in chick diet, whereas sample from eleven nests in Ukraine dominated by spiders (43%) and lepidopterans (27%); diet of nestlings also includes, in lesser amounts, orthopterans, hemipterans, dipterans, hymenopterans, coleopterans, molluscs, and sometimes fruits. Considered less frugivorous than are more specialized congeners such as C. borin or Sylvia atricapilla, but berries exploited on large scale during autumn migration; in Spain and Portugal consumes extensively fruits of the genera Pistacia, Ficus, Rubus, Daphne, Phillyrea and Rhamnus (up to 75% of total items consumed in a sample from Spain). Other fruits frequently taken are Ribes, Hippophae, Osyris, Smilax, etc. Berries recorded in diet also during winter or on spring passage in Africa; in addition to insects, Salvadora and Maerua berries taken regularly, and in S Africa small fruits of Grewia; in Baluchistan (Pakistan) seen to feed on Elaeagnus angustifolia berries. In Nigeria, stomachs of 97 birds in spring contained 80% (dry weight) fruits and flowers of Salvadora, 15% midges (Tanytarsus) and 5% other insects. During spring migration recorded as feeding on nectar at stopover sites (Italian islands), and in Denmark samples of stomach contents from two successive years (627 items and 524 items, respectively) contained 27–29% larval lepidopterans, 19–20% curculionid beetles and 12–14% nymphal psyllids (Psyllidae). Seeds also recorded in diet. Forages mainly in bushes and other low vegetation, taking items mostly from leaves and twigs; occasionally catches insects in flight or on ground. Of 148 prey items seen captured in study in Kenya, 85% were obtained from leaves and 13% from twigs or stems; further, of 40 observed feeding attempts, 80% were inside bushes or trees at low height.
Sounds and Vocal Behavior
Song , from dense cover, open perch or in flight, variable and complex (up to three types of song and 25 calls distinguished), a rapid, dry and somewhat musical warble described as “cheechiwee-cheechiweechoo-chiwichoo”, usually alternating high and low notes and imitations, commonly of congeners, in final strophes; lasts from c. 1·5 seconds (perched) to up to 10 seconds (in flight). Common contact call a nasal “wheet-wheet-wheet”, used also as alarm, and in cases of conflict accompanied by “dschid” and similar but more guttural notes, varying according to degree of excitement; usual alarm a harsh buzzy “dchaer” or “charr” and also prolonged “dschrááid”; occasionally utters a “tek” note, also a rattling, sometimes quite extended “drrt”.
Breeding
Mostly Apr–Jul, starting later in N Europe, and precise timing varies markedly with altitude; generally one clutch per season, but two broods not rare in S of range. Mostly monogamous, but some males bigamous and hold more than one territory; males arrive in breeding areas some days earlier than females and initially occupy larger territories of c. 4 ha (where mating occurs), subsequent breeding territory considerably smaller. Male has conspicuous song flight, rises for up to 10 m, holding tail spread and beating wings in characteristic butterfly-like manner, before dropping back to initial perch or different one. Male constructs several “cock nests”, female either selects and completes one of these or builds new nest in different site; a fairly deep cup constructed from grass, leaves, rootlets, spider cocoons and hair, concealed low down in bush or tall grass, in Britain mostly in bramble (Rubus fruticosus), nettles (Urtica) or grasses; strong fidelity to breeding area in successive years. Clutch 4–5 eggs, 2–3 replacements laid if clutches lost; incubation by both sexes, period 9–14 days; chicks fed by both parents, nestling period 10–12 days; young independent 15–20 days after fledging ; when male bigamous, secondary brood normally reared by female alone, but exceptions reported and even cases of two replacement clutches attended simultaneously by male and both females. Breeding success in Britain estimated at 58·6% (68·9% of clutches hatched, 85% of hatchlings fledged), mean 2·7 young fledged per nest (including unsuccessful nests), main causes of failure include desertion (31·6%), mowing (7·9%), unseasonable weather (4·6%) and parasitism by Common Cuckoo (Cuculus canorus) (1·3%); in Sweden, estimated success 73% (81·1% hatched, of which 90·2% fledged). Oldest recorded individual 8 years; causes of mortality of ringed individuals in NW Europe are domestic predators 19%, human-related (accidental) 49%, human-related (deliberate) 25%, other 7%.
Conservation Status
Not globally threatened. Common. Recent estimates put European population at more than 14,000,000 breeding pairs, and considered largely stable in period 1970–1990, with possible small increase during 1990–2000. Favourable habitats can hold up to 15 pairs/10 ha. Densities during breeding difficult to estimate, as territory size varies throughout season and unmated males also defend temporary territories; in large areas pairs aggregated in some places, whereas other areas unoccupied, thus densities of 0·5–5 pairs/km² recorded in Germany and Switzerland and 0·1–0·5 pairs/km² in Netherlands. Density, however, highly variable and locally can reach 100 pairs/km² in favourable habitat (e.g. Switzerland), and even 250–800 pairs/km² in Ukraine; on non-breeding grounds, densities of up to 0·9–3 birds/ha reported in Botswana. Marked annual fluctuations; strong decline (c. 50–90%) in populations of C & W Europe was caused by continuous drought in Sahel zone of W Africa during period 1968–1975 (especially 1968 and 1969). In addition to lack of rainfall, changes in the Sahel resulting from human pressure have led to increased desertification; in Europe, changes in land use through intensification of agriculture, combined with destruction of hedgerows and bushes, reduce the amount of available habitat suitable for breeding. Although populations recovered since the dramatic decline in 1969, densities have rarely reached same levels as those recorded before.
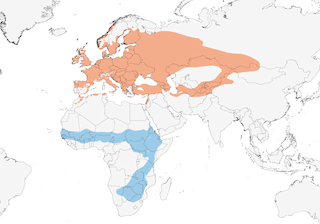
- Year-round
- Migration
- Breeding
- Non-Breeding

















