











Turquoise-browed Motmot Eumomota superciliosa Scientific name definitions
Text last updated December 18, 2012
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Catalan | motmot cellut |
| Czech | momot skořicovobřichý |
| Dutch | Wenbrauwmotmot |
| English | Turquoise-browed Motmot |
| English (United States) | Turquoise-browed Motmot |
| French | Motmot à sourcils bleus |
| French (France) | Motmot à sourcils bleus |
| German | Brauenmotmot |
| Icelandic | Grænpendill |
| Japanese | アオマユハチクイモドキ |
| Norwegian | turkisbrynmotmot |
| Polish | piłodziób czarnogardły |
| Russian | Синебровый момот |
| Serbian | Momot sa tirkiznom obrvom |
| Slovak | momot čiernohrdlý |
| Spanish | Momoto Cejiazul |
| Spanish (Costa Rica) | Momoto Cejiceleste |
| Spanish (Honduras) | Taragón Ceja Turquesa |
| Spanish (Mexico) | Momoto Cejas Azules |
| Spanish (Spain) | Momoto cejiazul |
| Swedish | turkosbrynad motmot |
| Turkish | Turkuaz Kaşlı Motmot |
| Ukrainian | Момот рудочеревий |
Eumomota superciliosa (Sandbach, 1837)
Definitions
- EUMOMOTA
- superciliosa / superciliosum / superciliosus
The Key to Scientific Names
Legend Overview
Originally Appeared in

Introduction
The Turquoise-browed Motmot is surely one of the most flamboyantly plumaged of a remarkable family, and it is sufficiently distinctive from other motmots to be afforded its own genus. Given its relatively small range, in the Yucatán Peninsula of eastern Mexico and northernmost Guatemala, and on the Pacific slope of Middle America, from southern Mexico south to northwest Costa Rica, it might seem remarkable that as many as seven subspecies are generally recognized. Virtually all of them have largely olive-green underparts with a cinnamon or rufous belly, and a darker reddish patch on the mantle, but the most eye-catching features are the turquoise flash in the flight feathers, the similarly colored eyebrow contrasting with the black mask, and the long graduated bright blue tail, which has the central feathers twice as long as the rest. Although they are reduced to mere shafts over more than half their length, they are ‘capped’ by black-tipped turquoise rackets. The dark bill is relatively long and slightly decurved.
Field Identification
33–38 cm; 44–74 g, means 67·6 g (male) and 66 g (female) (1). Tail most extremely graduated of family, central rectrices about twice as long as next pair, with bare shafts very long (c. 75 mm), flag-like racquet tip nearly twice as wide as basal part of vane, the two flanges decurved, dropping away from shaft on each side; bill narrow, cutting edges of both mandibles finely serrated along most of length. Racqueted shape of tail attained when weakly attached barbs deliberately shed, with barbs along wire weak due to combination of reduced structural attachment and lack of structurally enhancing melanin in proximal rami (2); unlike many avian species with “pin” tails, E. superciliosa has the distal portion of the rachis reinforced (3). Both sexes in nominate race conspicuous pale, slightly iridescent, turquoise-blue eyebrow, black facial mask with turquoise line below; mainly green above, with chestnut-rufous patch on mantle; flight-feathers turquoise-blue, black-tipped; tail-racquets sharply divided into blue basal half, black distal half; throat and breast olive-green with shaggy black medial stripe edged with turquoise, belly cinnamon; bill black; legs and feet grey. Male larger than female in most body-size characters, including those of the tail (1), with the latter probably functioning as a sexual signal (4). Furthermore, recent detailed research has revealed that tail colour in both sexes has similar spectral shape, with significant, albeit moderate, dichromatism, males being brighter than females and having marginally greater blue-green saturation, while length of feather grown per day is positively related to overall feather brightness in males, suggesting that tail colour potentially imparts information about individual quality during mate choice (5). Distinguished from other momotids by exceptionally long bare tail shafts, blue eyebrow, pattern of throat and breast. Juvenile duller, little or no rufous on back, little of black-and-turquoise stripe below. Races vary mainly in darkness of plumage: bipartita darker, greenish-olive breast and cinnamon-rufous flanks and belly sharply demarcated; sylvestris like previous but generally darker; euroaustris darker turquoise-blue eyebrow, darker wings and tail more purplish-blue; vanrossemi like nominate but brighter green, tawny suffusion reduced; apiaster with green paler, less tawny suffusion; <em>australis</em> distinctive, being small and pale.
Systematics History
Editor's Note: This article requires further editing work to merge existing content into the appropriate Subspecies sections. Please bear with us while this update takes place.
Highly distinctive, with no very close relatives. Geographical variation within disjunct range is probably complex, but general pattern of darkest forms in more humid areas, tawnier forms coastal, greener ones isolated in interior, and those in far S distinctly pale; further study required. Seven subspecies currently recognized.Subspecies
Eumomota superciliosa bipartita Scientific name definitions

Distribution
Eumomota superciliosa bipartita Ridgway, 1912
Definitions
- EUMOMOTA
- superciliosa / superciliosum / superciliosus
- bipartita / bipartitus
The Key to Scientific Names
Legend Overview
Eumomota superciliosa superciliosa Scientific name definitions

Distribution
Eumomota superciliosa superciliosa (Sandbach, 1837)
Definitions
- EUMOMOTA
- superciliosa / superciliosum / superciliosus
The Key to Scientific Names
Legend Overview
Eumomota superciliosa vanrossemi Scientific name definitions
Distribution
Eumomota superciliosa vanrossemi Griscom, 1929
Definitions
- EUMOMOTA
- superciliosa / superciliosum / superciliosus
- vanrossemi
The Key to Scientific Names
Legend Overview
Eumomota superciliosa sylvestris Scientific name definitions
Distribution
Eumomota superciliosa sylvestris Carriker & Meyer de Schauensee, 1935
Definitions
- EUMOMOTA
- superciliosa / superciliosum / superciliosus
- sylvestris
The Key to Scientific Names
Legend Overview
Eumomota superciliosa apiaster Scientific name definitions
Distribution
Eumomota superciliosa apiaster (Lesson, 1842)
Definitions
- EUMOMOTA
- superciliosa / superciliosum / superciliosus
- apiaster
- Apiaster
The Key to Scientific Names
Legend Overview
Eumomota superciliosa euroaustris Scientific name definitions
Distribution
Eumomota superciliosa euroaustris Griscom, 1929
Definitions
- EUMOMOTA
- superciliosa / superciliosum / superciliosus
- euroaustris
The Key to Scientific Names
Legend Overview
Eumomota superciliosa australis Scientific name definitions

Distribution
Eumomota superciliosa australis Bangs, 1906
Definitions
- EUMOMOTA
- superciliosa / superciliosum / superciliosus
- australe / australis
The Key to Scientific Names
Legend Overview
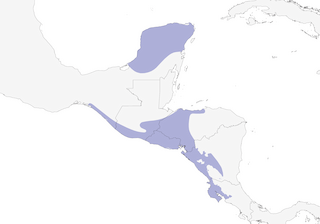
Distribution
Editor's Note: Additional distribution information for this taxon can be found in the 'Subspecies' article above. In the future we will develop a range-wide distribution article.
Habitat
Deciduous woodland and woodland edge, evergreen gallery forest, low scrubby thickets, and semi-open areas with scattered trees; also locally plantations and gardens; to 1400 m.
Movement
Returns to Yucatán breeding colonies in late Feb or Mar, a few weeks before start of nesting; extent of movement unknown, probably not great and speculated to only move as far as moist forests of C Yucatán (1). Some seasonal movement probable also in Costa Rica , where more arid habitats occupied mainly in wet season.
Diet and Foraging
Insects , spiders, millipedes, small molluscs, earthworms, and small lizards and snakes; also fruit. Coleoptera, Orthoptera and Hymenoptera are especially important dietary constituents, whereas reptiles constitute just 0·9% by volume (6). Fruit also in small quantities, e.g. 3·4% by volume of stomach contents in Yucatán; species recorded taken include Stemmadenia, Ficus, Bursera, Ehretia and Guaiacum. Observed feeding on flying insects around artificial streetlights almost one hour after sunset, in El Salvador (7). Animal food seized in sudden dart to vegetation or ground, or in aerial sally; butterflies, bees, dragonflies and smaller insects also taken in flight. Larger prey is struck up to eight times against perch prior to consumption (7).
Sounds and Vocal Behavior
Like other motmots, gives a hollow, nasal, ringing "wohh" or "owhh", given at rate of one per 3·5–4·5 sec and higher and less resonant than Electron motmots, with varied, also hollow, but slightly hoarse, clucking calls, including a rhythmic "k'wok k'wok..." and a longer "k'wok t'k'wok t'k'wok t'k'wok..." (the latter perhaps in duet), have also been recorded (8).
Breeding
Lays second week May to mid-Jul (peak first half Jun) in Mexico; Apr–May in Guatemala and Honduras; late Mar to mid-Apr in NW Costa Rica (9); may re-lay after early nest failure. Contrary to some predictions, no evidence of assortative mating for tail components, body size or phenotypic condition, in Yucatán population, although same study uncovered some evidence that birds >2 years old select partners of a similar age and phenotypic condition (1). Pair-bond maintained over successive years, and individuals faithful to nest-sites; either solitary or colonial, depending on availability of suitable nest-sites, some colonies number over 100 pairs, but in the Yucatán colonies range in size from 2–60 pairs, with 10–20 pairs most common (1). Nest typically in burrow excavated in earth bank or low cliff; burrow often curved, total length very variable, 40–244 cm (mean 130 cm) (1), ending in enlarged nest-chamber c. 20–23 cm wide and c. 10 cm high; in Yucatán, Mexico, also uses beam-socket and ventilation holes of archaeological ruins, sinkholes and freshwater wells (1). Clutch 2–6 eggs, most often four, laid at intervals of c. 48 hours; incubation 18–20 days; fledging 24–31 days. In Yucatán archaeological sites, 36% of all nesting attempts failed owing to predation, probably by rat-snake (Elaphe phaescens).
Conservation Status
Not globally threatened. One of the most abundant motmots, common to fairly common throughout range. Well adapted to secondary vegetation and semi-open country in areas much altered by man; also able to exploit nesting opportunities created by man, where breeding adults become conditioned to presence of human visitors. The national bird of El Salvador (10).
- Year-round
- Migration
- Breeding
- Non-Breeding

















