











Yellowhammer Emberiza citrinella Scientific name definitions
Text last updated March 1, 2013
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Albanian | Cërla verdhashe |
| Arabic | درسة ذهبية |
| Armenian | Սովորական դրախտապան |
| Asturian | Escribana mederiza |
| Azerbaijani | Adi vələmirquşu |
| Basque | Berdantza horia |
| Bulgarian | Жълта овесарка |
| Catalan | verderola |
| Chinese (SIM) | 黄鹀 |
| Croatian | žuta strnadica |
| Czech | strnad obecný |
| Danish | Gulspurv |
| Dutch | Geelgors |
| English | Yellowhammer |
| English (United States) | Yellowhammer |
| Faroese | Gulspurvur |
| Finnish | keltasirkku |
| French | Bruant jaune |
| French (France) | Bruant jaune |
| Galician | Escribidor amarelo |
| German | Goldammer |
| Greek | Χρυσοτσίχλονο |
| Hebrew | גיבתון צהוב |
| Hungarian | Citromsármány |
| Icelandic | Gultittlingur |
| Italian | Zigolo giallo |
| Japanese | キアオジ |
| Korean | 노랑멧새 |
| Latvian | Dzeltenā stērste |
| Lithuanian | Geltonoji starta |
| Mongolian | Алтан хөмрөг |
| Norwegian | gulspurv |
| Persian | زردپره لیمویی |
| Polish | trznadel |
| Portuguese (Portugal) | Escrevedeira-amarela |
| Romanian | Presură galbenă |
| Russian | Обыкновенная овсянка |
| Serbian | Strnadica žutovoljka |
| Slovak | strnádka obyčajná |
| Slovenian | Rumeni strnad |
| Spanish | Escribano Cerillo |
| Spanish (Spain) | Escribano cerillo |
| Swedish | gulsparv |
| Turkish | Sarı Çinte |
| Ukrainian | Вівсянка звичайна |
Emberiza citrinella Linnaeus, 1758
Definitions
- EMBERIZA
- citrinella
- Citrinella
The Key to Scientific Names
Legend Overview
Originally Appeared in

Field Identification
16–16·5 cm; 20–36·5 g. Rather large bunting. Male nominate race in fresh plumage (Sept–Nov) has yellow head with broad black and olive lateral crownstripe (most of crown feathers yellow, with relatively well-demarcated tiny dark greyish-green spot on tip), more or less diffuse dark border around ear-coverts (appearing broken at upper rear corner); warm brown upperparts streaked blackish, much less streaked on rufous-chestnut rump and uppertail-coverts; edges of primaries with yellow tones, and white in outer rectrices; throat pale yellowish, streaky dark malar stripe; below, yellowish with blackish streaks, especially on breast and rear flanks, with olive chestband and elements of rufous on side of breast and upper flanks; iris dark brown; bill blue-grey, slightly paler below; legs flesh-brown. Male in worn plumage (May–Aug) has head extensively yellow, dark pattern sometimes restricted to narrow blackish streaks above supercilium, often very faint above eye, sometimes restricted to small dark area at base of bill and at rear, and narrow border around ear-coverts, with breast more prominently rufous; supercilium lemon-yellow behind eye, anteriorly more greenish-yellow (approaching colour of lores), dark greenish eyestripe curving down around rear ear-coverts to lower rear corner, malar stripe usually absent; nape a mixture of yellow and greenish-grey, upperparts more richly coloured than in fresh plumage. Female in fresh plumage is in general more variable than male, with pattern similar, but less prominent and brilliant colours above, and slightly paler yellow below, much broader olive on crown, ear-covert markings, malar stripe and throat marks, olive nape and band around lower neck and shoulder, breaking up on side of breast; in the hand, differs from male in having greyish crown tips usually more extensive, with more and longer prominent dark shaft streak and reduced amount of yellow. Female in worn plumage shows less extensive yellow on head and more streaked crown, also grey-green head side with no yellow, otherwise similar in pattern to male, but in general with rather dull ground colours above and paler yellow below, where greyish-black streaking on breast and flanks more marked, and throat pale yellow; in the hand, crown feathers have the greyish tip usually more extensive, with more and longer prominent dark shaft streak and reduced amount of yellow, compared with male. First winter is rather less distinctive, with less yellow on head, which usually almost wholly brown, lacking obvious supercilium, and with diffuse streaks on crown, and buff-white underparts with only some faint yellow tinge; first-winter male plumage similar to adult female, except for brighter yellow and chestnut tracts and reduced size of marks on head; first-winter female duller than adult in similar plumage, but still lacking olive on crown and nape. Juvenile is variable, but has pattern of more intense streaking on underparts and duller colours than average adult female, with indistinct supercilium, and broader blackish streaks on mantle and scapulars. Race <em>caliginosa</em> male is slightly darker, more richly coloured and with more streaks than nominate; <em>erythrogenys</em> is paler and less well streaked than male nominate, with hindneck and shoulders greyish (rather than greenish), and with yellow pigments and chestnut on flanks and breast richer.
Systematics History
Editor's Note: This article requires further editing work to merge existing content into the appropriate Subspecies sections. Please bear with us while this update takes place.
Sometimes considered conspecific with E. leucocephalos, but clear differences in plumage, as well as in behaviour and osteology, support treatment as separate species. The two hybridize extensively in W & C Siberia (across c. 3000 km in taiga and forest-steppe zone from R Ural E to L Baikal), and intensity of interbreeding is increasing, since each species is expanding its breeding range across/towards that of the other; this type of interbreeding has been described as intermediate between occasional hybridization (not influencing gene pools of parent species) and introgressive hybridization (resulting sometimes in local fusion of species). An independent analysis of phylogeny supports recognition of two independent species, although cytochrome b sequences very similar, giving strong indication that they are sister-species; sharing of haplotypes even in very distant locations seems to be result of introgression of mtDNA. Independent alternative study also argues that both species are older than their closest relatives and that mtDNA has recently introgressed between them, likely as result of selective sweep. Has been suggested that race erythrogenys is product of hybridization with E. leucocephalos. Nominate race intergrades with erythrogenys in broad zone from Baltic republics, W European Russia and W Belarus S to E Hungary, E & S Balkans and Greece. Three subspecies recognized.Subspecies
Introduced in New Zealand region.
Emberiza citrinella caliginosa Scientific name definitions
Distribution
Emberiza citrinella caliginosa Clancey, 1940
Definitions
- EMBERIZA
- citrinella
- Citrinella
- caliginosa / caliginosus
The Key to Scientific Names
Legend Overview
Emberiza citrinella citrinella Scientific name definitions
Distribution
Emberiza citrinella citrinella Linnaeus, 1758
Definitions
- EMBERIZA
- citrinella
- Citrinella
The Key to Scientific Names
Legend Overview
Emberiza citrinella erythrogenys Scientific name definitions
Distribution
Emberiza citrinella erythrogenys Brehm, 1855
Definitions
- EMBERIZA
- citrinella
- Citrinella
- ERYTHROGENYS
- erythrogenys
The Key to Scientific Names
Legend Overview
Hybridization
Hybrid Records and Media Contributed to eBird
-
Yellowhammer x Pine Bunting (hybrid) Emberiza citrinella x leucocephalos
Distribution
Editor's Note: Additional distribution information for this taxon can be found in the 'Subspecies' article above. In the future we will develop a range-wide distribution article.
Habitat
Breeds mainly in open woodlands, scrub, heathland, cultivations and similar habitats, also riparian forest and orchards, avoiding dense forest. Most abundant between 600 m and 900 m; in S of range a rather montane species, inhabiting clearings in forest to 2000 m, especially in areas where cereals are cultivated on higher slopes. In Ukraine and Russia extends into steppe oakwood (Quercus); at margin of forest tundra and true tundra in N of range in E. In winter mostly in agricultural land , often stubble fields, unharvested cereal fields, pastures, margins of fields (where weed seeds may be found), orchards, edges of forest and on waste ground. Roosts in trees or bushes, sometimes in reeds.
Movement
Sedentary and partially migratory. Only extreme N part of range completely vacated during winter. In W part of range autumn movements Sept–Nov, arriving regularly in winter in areas of coastal and inland Spain (where absent as a breeding species), as well as in coastal areas of Italy (where also not breeding), and similar pattern in parts of Balkans, most of Black Sea region and Mediterranean in Turkey, S to N Israel; in severe winters extends S to S Spain, N Africa, Cyprus and United Arab Emirates. E populations dispersive and migratory, wintering S mainly to Transcaucasia, N Iran, S Kazakhstan and N Mongolia, minority resident in W Siberia.
Diet and Foraging
Mostly seeds; during breeding season, at least, also invertebrates. Recorded foods include seeds and other parts of spruce (Picea), pine (Pinus), beech (Fagus), grape (Vitis), mistletoe (Viscum), and several other plant species, such as those of genera Urtica, Rumex, Polygonum, Chenopodium, Stellaria, Cerastium, Rubus, Vicia, Trifolium, Empetrum, Myosotis, Coryza, Taraxacum, Centaurea, Sonchus, Achillea, Helianthus and Plantago, and several grasses (Poaceae), especially cereals. Several studies in UK farmland areas during autumn–winter reveal high consumption of cereal grain (wheat and oats constantly preferred over barley), as well as a few common weed species, taken in proportion to their availability and size. During breeding season, shift towards invertebrates is related to feeding of nestlings. Takes wide array of invertebrates, which include springtails (Collembola), mayflies (Ephemeroptera), grasshoppers and crickets (Orthoptera), cockroaches (Blattodea), earwigs (Dermaptera), bugs (Hemiptera), lacewings (Neuroptera), adult and larval butterflies and moths (Lepidoptera), caddis flies (Trichoptera), flies (Diptera), Hymenoptera, beetles (Coleoptera), spiders (Araneae), woodlice (Isopoda), millipedes (Diplopoda), earthworms (Lumbricidae), and snails (Gastropoda). Forages almost exclusively on ground , but recorded also as clinging from tall herbs, especially during snowfalls. During breeding season, different techniques for catching insects recorded, including brief pursuits of flying insects, also jumping from ground, and even briefly hovering from branches on trees. Outside breeding season, feeds in flocks of variable size, usually of the order of tens of individuals, occasionally hundreds, rarely more (flocks of up to 3000 individuals recorded in Germany); often with mixed-species flocks containing other buntings and finches (Fringillidae). Roosts of variable size, normally of several tens or hundreds of individuals, sometimes up to 1000, usually also with other species such as E. calandra and some finches.
Sounds and Vocal Behavior
Song , generally from top of tree, bush or wire, consists usually of a rather constant series of 5–12 similar notes, usually ending in drawn-out descending note of different pitch, e.g. “zi-zi-zi-zi-zi-zi-zi-ziiiiiiiii”, penultimate note usually a little higher before the descent of lower final note; geographical dialects occur. In analysis of territorial songs of different males four different structures distinguished, with frequency ranges of elements more constant across individuals than were the ranges of the notes, and among the latter the low-frequency element was nearly twice as conserved as the high-frequency element. Call notes characteristic, the commonest a “stif”, as well as an array of subtle variations of similar clicking calls, e.g. “pt...pt”, also “trp”, and a fine “tsii” described as alarm call.
Breeding
Normally begins in Apr, later at higher latitudes, with late broods started in Sept; most first broods started May in W & C Europe; normally two broods, sometimes three. Nest built by female, sometimes joined by male on movements to/from site, material gathered from immediate surroundings of (normally within maximum radius of 50 m); composed of dry grass, plant stems, leaves and some moss, lined with rootlets, fine grass and sometimes animal hair, external diameter 11·5–13 cm, internal diameter 5·5–7 cm, depth of cup 4–4·5 cm, placed on or near ground, well hidden among grass or herbage, typically against base of herb, bush or small tree, or inside well-protected thorn bush such as Rubus. Clutch 3–5 eggs, white, tinged bluish, greyish or purplish, usually some fine faint pale violet or purple spotting, rather variable in colour; incubation usually by female alone, period 12–14 days; chicks brooded entirely by female, male delivers food, nestling period 11–13 days. Breeding success in Britain was 58% of 244 hatched eggs in complete clutches, hatching success increasing from May to later in season; 44% of clutches produced fledged young at a locality in Britain, 52% did so at a German site; nests frequently preyed on by rodents and by corvids including crows (Corvus), Eurasian Jay (Garrulus glandarius) and Common Magpie (Pica pica). Predation accounted for 64% of nest failures; in one UK study examining variation in habitat selection and breeding success, territories found to be associated with hedgerows, vegetated ditches and wide uncultivated grassy margins around fields, avoiding pastures and silage leys, breeding starting slightly earlier on organic farms than on intensively managed ones, but with no differences in breeding success; another study found relationship between mixed farming and improvement of breeding success (abandonment of mixed-farming practice adversely affected success).
Conservation Status
Not globally threatened (Least Concern). Locally common to very common; scarce in some areas. Introduced in New Zealand between 1862 and early 1870s (1), and now widespread and common throughout the islands; has also reached other islands in that region, e.g. Kermadecs, Lord Howe I. In Europe (including European Russia), breeding population estimated to number 18,000,000–31,000,000 breeding pairs, and considered stable during 1970–1990, equating to c. 54,000,000–93,000,000 individuals. Europe forms 50–74% of global range, and a very preliminary estimate of global population is 73,000,000–186,000,000 individuals; further estimates based on more reliable data, especially for Russia, are needed. Although declines recorded in Fennoscandia, Latvia and W Europe (Ireland, UK , Belgium, Netherlands), Austria and Italy during 1990–2000, most C & E European populations (including sizeable ones in Germany, Czech Republic, Poland and Ukraine) were stable; Russian trend not known. Changing land use and farming practices affect populations densities locally.
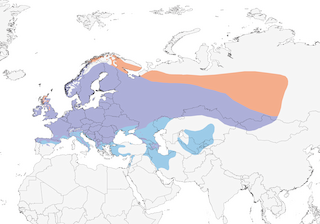
- Year-round
- Migration
- Breeding
- Non-Breeding



















