










Streaked Shearwater Calonectris leucomelas Scientific name definitions
- NT Near Threatened
- Names (36)
- Monotypic
Text last updated March 17, 2017
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Afrikaans | Gestreepte Pylstormvoël |
| Arabic | جلم ماء مخطط |
| Catalan | baldriga ratllada |
| Chinese | 大水薙鳥 |
| Chinese (Hong Kong SAR China) | 白額鸌 |
| Chinese (SIM) | 白额鹱 |
| Czech | buřňák světlečelý |
| Danish | Hvidmasket Skråpe |
| Dutch | Gestreepte Pijlstormvogel |
| English | Streaked Shearwater |
| English (United States) | Streaked Shearwater |
| French | Puffin leucomèle |
| French (France) | Puffin leucomèle |
| German | Weißgesicht-Sturmtaucher |
| Greek | Γραμμωτός Αρτέμης |
| Hebrew | יסעור לבן-מצח |
| Hungarian | Csíkos vészmadár |
| Icelandic | Fjöruskrofa |
| Indonesian | Penggunting-laut belang |
| Japanese | オオミズナギドリ |
| Korean | 슴새 |
| Lithuanian | Dryžagalvė audronaša |
| Malayalam | വരയൻ തിരവെട്ടി |
| Norwegian | hvitpannelire |
| Polish | burzyk kreskowany |
| Portuguese (Portugal) | Cagarra-do-pacífico |
| Romanian | Ielcovan vărgat |
| Russian | Пестролицый буревестник |
| Serbian | Beloliki zovoj |
| Slovak | víchrovník jarabý |
| Spanish | Pardela Canosa |
| Spanish (Spain) | Pardela canosa |
| Swedish | vithuvad lira |
| Thai | นกจมูกหลอดลาย |
| Turkish | Ak Yüzlü Yelkovan |
| Ukrainian | Буревісник тихоокеанський |
Calonectris leucomelas (Temminck, 1836)
Definitions
- CALONECTRIS
- leucomelas
The Key to Scientific Names
Legend Overview
Originally Appeared in

Introduction
In many ways the East Asian counterpart of the European-distributed Cory’s Shearwater (Calonectris borealis) complex, the Streaked Shearwater breeds in northeast Japan, and on the Izu Islands and Ryukyu Islands, as well as on islands off the Russian Far East, eastern China and the Korean Peninsula. In the non-breeding season at-sea dispersal is extensive, although a comparatively small percentage of the well in excess of one million-strong population remain in Japanese waters at this time, most migrate south towards Australia and large flocks occur off New Guinea, with the southernmost record in New Zealand, while some move east to Hawaii and and the Pacific coast of the United States, as well as west into the Indian Ocean, where it has reached as far afield as South Africa and the head of the Red Sea at Eilat, in southern Israel. The latter records, in the early 1990s, permitted the observers to practice separating this species from the much more regular Cory’s Shearwater, versus which it differs in having a more wedge-shaped tail, and especially by its pale face, contrasting dark nape, and relatively slenderer bill.
Field Identification
Large, distinctive, brownish shearwater with mostly clean white underparts, mottled head (due to variable white spots and streaks on face and crown), and rather large, pale bill, with variable dark streaks on the underside of the primary-coverts.
Similar Species
Recalls Cory's Shearwater (<em>C. diomedea</em>) in size, flight action and upperparts colour, usually less so on underwing , but separated by more wedge-shaped tail, and especially by pale face and contrasting dark nape, while bill is relatively slender, similar to that of Cape Verde Shearwater (C. edwardsii). Same features distinguish it from large Pterodroma. From other shearwaters by pale face and bill, and by lazy flight on rather bowed wings.
Plumages
Juvenile
As adult.
Adult
Head variably mottled white and blackish brown, the white usually predominating around eye, on forehead and foreface, the dark markings being larger and more solidly dark on nape, rest of upperparts gray-brown, pale feather fringes affording scaly appearance to mantle, scapulars, rump and uppertail-coverts, the latter often with obvious white tips that can look like narrow white horseshoe. Most of upperwing slightly darker brown, darkest on primary-coverts and remiges, tertials and lowermost scapulars also dark, often giving upperparts diffuse dark M pattern. Uppertail blackish brown; underwing is dark grey on exposed remiges forming dark trailing edge much broader at wingtip, otherwise white except dark spot on some of outer greater primary-coverts, median primary-coverts usually have mostly dark outer webs forming broken dark crescent on centre of outer wing, narrow dark streaks on leading edge in carpal region and on lesser coverts forming weak dark diagonal band on underwing, but dark markings variable and some birds have nearly all-white underwing-coverts. Sides of neck to sides of chest brownish with blurry pale scaling, rest of underparts white. Sexes alike in plumage, although males have significantly deeper and longer bills, longer heads, longer tarsi, greater wingspans and greater wing areas than females, but overall wing and tail lengths, and aspect ratios do not differ between sexes, while the species shows greater degree of sexual dimorphism in bill size than other Calonectris species (1).
Molts
Poorly known. Adults are probably molting their wing feathers mainly between November and March; and first-years between September and January (2).
Bare Parts
Bill
Pale horn to grey, with some dusky gray separating the tip from the rest of the bill (often looks dark-tipped in field) (3).
Iris
Blackish brown (3).
Legs and Feet
Pink, but can have dusky-gray outer toes (3).
Measurements
Linear Measurements
Overall length 45–52 cm (4).
Linear measurements (in mm, with means, standard deviation, and sample sizes in parentheses, taken during the incubation or chick-rearing period at eight breeding colonies in Japan, most of them around Honshu 5):
| Funakoshi-Ohshima | Sanganjima | Awashima | Kanmurijima | Mikurajima | Uwashima | Danjo-gunto | Nakanokamishima | |
| Males | ||||||||
| Wing length | (312–345 (325 ± 8, n = 14) | (310–340 (324 ± 7, n = 54) | (308–343 (322 ± 8, n = 47) | (309–331 (318 ± 7, n = 14) | (307–326 (316 ± 5, n = 32) | (305–342 (322 ± 9, n = 11) | (316–326 (322 ± 4, n = 7) | (273–323 (299 ± 8, n = 43) |
| Tarsus length | (51.5–56.7 (53.9 ± 1.7, n = 14) | (50.0–55.7 (53.7 ± 1.2, n = 54) | (50.6–56.0 (53.2 ± 1.3, n = 47) | (50.4–54.1 (52.5 ± 1.2, n = 14) | (49.6–55.0 (52.2 ± 1.2, n = 32) | (50.6–55.2 (53.3 ± 1.5, n = 11) | (48.9–55.6 (51.7 ± 2.3, n = 7) | (47.6–54.1 (50.4 ± 1.4, n = 43) |
| Bill length | (49.4–54.4 (51.7 ± 1.6, n = 14) | (48.7–54.2 (51.6 ± 1.5, n = 54) | (46.4–54.2 (51.3 ± 1.6, n = 47) | (48.0–54.0 (50.9 ± 1.7, n = 14) | (46.0–54.3 (50.3 ± 1.8, n = 32) | (47.1–51.8 (50.4 ± 1.6, n = 11) | (47.6–51.8 (49.6 ± 1.8, n = 7) | (44.8–52.9 (48.4 ± 1.5, n = 43) |
| Bill depth | (11.6–13.5 (12.5 ± 0.6, n = 14) | (11.9–15.1 (13.3 ± 0.6, n = 54) | (11.0–15.1 (12.9 ± 0.8, n = 47) | (11.5–13.8 (12.6 ± 0.6, n = 14) | (11.1–14.0 (12.4 ± 0.6, n = 32) | (9.3–13.5 (12.2 ± 1.2, n = 11) | (11.8–14.1 (12.9 ± 0.8, n = 7) | (10.8–13.0 (11.9 ± 0.6, n = 43) |
| Females | ||||||||
| Wing length | 301–323 (313 ± 7, n = 10) | (305–331 (317 ± 6, n = 52) | (300–326 (315 ± 8, n = 40) | (294–318 (308 ± 5, n = 22) | (297–325 (311 ± 7, n = 26) | (293–329 (311 ± 8, n = 23) | (305–323 (314 ± 5, n =13) | (271–303 (290 ± 7, n = 46) |
| Tarsus length | (49.2–53.7 (51.6 ± 1.5, n = 10) | (48.3–54.3 (51.6 ± 1.3, n = 52) | (47.4–54.0 (51.3 ± 1.4, n = 40) | (47.5–52.4 (50.6 ± 1.2, n = 22) | (48.2–53.0 (50.4 ± 1.3, n = 26) | (47.0–56.9 (50.9 ± 2.0, n = 23) | (47.9–53.3 (50.3 ± 1.2, n =13) | (45.4–50.1 (47.3 ± 1.3, n = 46) |
| Bill length | (46.9–50.5 (48.5 ± 1.1, n = 10) | (44.0–51.9 (48.2 ± 1.7, n = 52) | (45.8–51.2 (51.3 ± 1.6, n = 40) | (45.1–50.0 (47.4 ± 1.3, n = 22) | (44.5–50.0 (46.9 ± 1.5, n = 26) | (43.7–51.2 (47.7 ± 2.0, n = 23) | (45.9–49.6 (47.7 ± 1.4, n = 13) | (41.1–47.3 (44.4 ± 1.4, n = 46) |
| Bill depth | (10.1–11.8 (11.1 ± 0.5, n = 10) | (10.5–13.1 (11.6 ± 0.7, n = 52) | (10.3–13.0 (11.5 ± 0.6, n = 40) | (10.0–12.9 (11.3 ± 0.7, n = 22) | (7.9–12.1 (10.7 ± 1.0, n = 26) | (9.2–12.3 (11.2 ± 0.6, n = 23) | (11.0–13.0 (11.9 ± 0.6, n = 13) | (9.5–11.8 (10.8 ± 0.5, n = 46) |
Mass
348–710 g (5).
Wingspan
103–113 cm (4).
Systematics History
Sometimes placed in the genus Procellaria, or in Puffinus (6).
Geographic Variation
None.
Subspecies
Monotypic.
Distribution
Western Pacific Ocean and east Indian Ocean, breeding in northeast Japan, on the Izu Islands and Ryukyu Islands to the Pescadores Islands (off western Taiwan); also on islands off the southeast Russian Far East (3), eastern China and Korea.
Habitat
Marine and partly pelagic, also occurring over inshore waters (7). Typically breeds on offshore islands, occupying forested hills, e.g. of Machilus thunbergii (8).
Movement
During breeding season is generally concentrated in seas around Japan and off Korea, but overall range at this period extends east to c. 160°E (3) and south to the northern Philippines at 19°N (9,10). During incubation period, shearwaters breeding on the islands of Sangan and Mikura, in the northwest Pacific, foraged in the Kuroshio-Oyashio transition area, while those nesting on Awa Island mostly foraged in the Sea of Japan; although those from Mikura Island foraged in the northwest Pacific, the distance between their colony and foraging areas was greater (mean 645 km; range 546–756 km), compared to birds on Sangan (272 km; range 244–297 km) and Awa (228 km; range 75–518 km) (11,12). Those nesting on Awashima Island in the Sea of Japan move into the Pacific Ocean, via the Tsugaru Strait, to feed, perhaps especially in years when waters around the breeding grounds are warmer, but males appear to do more than females, apparently because their greater wing loadings creates higher energy demands that necessitate visiting better foraging areas (13). Small numbers remain in Japanese waters during winter (7), but most migrate south towards Australia between November and April (14,15,3): there are many ringing returns from Philippines; large flocks occur off New Guinea (16,17,18,19) and the species is regular elsewhere in Indonesian waters (3, 20,21), where mainly recorded from September, but also as early as May–June (22, 23); now known to be regular off northern Australia, with some moving along the west and east coasts to southern Australia, e.g. off New South Wales (24,3); and recently recorded (February 2006) as far southeast as New Zealand (25,26). A study using geolocators, attached to 48 breeding birds in 2006 and subsequently recovered from 38 birds the following year, confirmed that the species migrated from the seas around Japan to three wintering areas in the tropics, the seas off northern New Guinea, the Arafura Sea, and South China Sea (4,000 km, 5,400 km, and 3,500 km from their breeding colony, respectively), with most in the first-named area, in seas of low primary productivity but high skipjack tuna Katsuwonus pelamis abundance (27). Telemetric studies have demonstrated that juveniles from a colony in northern Honshu take a different route from adults during their first southward migration, initially moving south overland, crossing the island’s mountains, whereas adults travel over water (28). Vagrants have been recorded east to Hawaii (29) and the west coast of the USA (mid August to mid October), off Oregon (30) and California, with single inland records in California and Wyoming (31, 4), as well as west to southern India (32), Sri Lanka, the Maldives (33,34) (mainly September–February, but also July, in South Asian waters) (32, 35), South Africa (3) and, most exceptionally, at the head of the Red Sea at Eilat, on the Israel/Jordan border, in June to mid September 1992, and May–July 1993 (36 ,37, 38), as well in Egypt in April 2015 (EORC report).
Diet and Foraging
Mainly fish and squid. Birds rearing chicks on Mikura Island, Japan, made many short trips (< 2 days) and some long ones (4–10 days); during short trips, birds stayed in warmer Kuroshio and Kuroshio-Oyashio mixed water regions, and fed on Japanese anchovies (Engraulis japonicus), squid (Todarodes pacificus) and flying-fishes (Cypselurus hiraii), whereas during longer trips they visited the colder Oyashio region and fed on anchovies and Pacific saury Cololabis saira (39,12). Prey captured by surface-seizing; also makes shallow plunges (40). Study during chick-rearing period off Japan found that shearwaters made 50.3 landings per day (± 9·8, n = 12) and spent 44.8% (± 8, n = 12) of their time on the water (41), and another study reported that birds spent 76–96% of their time at sea flying, 4–24% landing on water and completed 0.5–17 dives per day, all of them shallow (< 6 m) (39). Often associates with other seabirds, especially other Procellariiformes, e.g. Wedge-tailed Shearwater Ardenna pacifica (42), Flesh-footed Shearwater A. carneipes (35) and Wilson’s Storm-petrel Oceanites oceanicus (22), but also in mixed-species feeding flocks containing frigatebirds (Fregata), boobies (Sula) (22), gulls (Larus spp.), and cormorants (Phalacrocorax spp.), among others (4), sometimes with cetaceans such as Short-finned Pilot Whales Globicephala macrorhynchus (22). Follows fishing boats; is attracted to anchovy crawls off Japan, and may feed on prey driven to the surface by predatory skipjack tuna Katsuwonus pelamis (42).
Sounds and Vocal Behavior
Vocal at colonies, where calls loudly and frequently both on ground and in flight, with evidence for some sexual dimorphism in vocalizations, with, for example, females uttering longer phrases (43,44,3) and lower-pitched calls (45).
Breeding
Starts March and ends in November, with pre-laying exodus of 13–17 days (females) or 4–7 days (males) (11), egg-laying commences in May and peaks in mid to late June (but sometimes does not start until July) (11), hatching generally peaks between 10 and 20 August, fledging from Octpber and departure continues through November, stragglers remaining into December (3). Strictly nocturnal at colonies (3), but presence there does not seem unduly influenced by ambient moonlight levels (46). Long-term study (1984–2010) on Kanmurijima Island, in the Sea of Japan, based on band recoveries, indicated high levels of site fidelity and natal philopatry; even after 20 years, birds returned to the vicinity of the initial release site and only very small percentages (0.7% and 0.2%) of returns were at other study sites (even then were on the same island) (47). Long-term monogamous, but recent study reported that extra-pair males sired 15% of 113 offspring during a study in 2014–2016, which percentage is among the highest recorded rates of extra-pair paternity among seabirds (48). Colonial; nests in burrows . Single white egg; incubation c. 45–64 days in five to six shifts (3, 11); fledging period c. 66–87 days (49). In Japan, duration of incubation shifts show significant inter-colony differences, but no sex-related differences; those on Sangan Island had shorter mean incubation shifts (5.6 days on average; range 3–8 days) than those on Mikura Island (7.2 days; range 4.8–10.7 days) and Awa Island (6.9 days; range 6.0–9.7 days) (11). Factors affecting breeding success are poorly known, but predators include feral cats (Felis catus) (49). No further information (50,51,52,3).
Conservation Status
Not globally threatened. Currently considered Near Threatened. Abundant and widespread, with total population of several million birds; well in excess of 1,000,000 pairs in Japan (7), of which largest concentration is in northern Izu Islands, most significantly 1,750,000–3,500,000 birds breeding on Mikura-jima (53), with fewer than 150 pairs in the Russian Far East, on Karamzin Island (54), while unknown numbers breed on islets off the Korea Peninsula, although recent surveys have demonstrated the existence of 19 confirmed colonies (one numbering almost 21,000 pairs) and four additional islands may also support nesting birds, and the species has probably been extirpated on three others (55). Human exploitation accounts for large numbers throughout the range (as many as 30,000 chicks used to be taken annually on Mikura-jima) (3), despite some protective laws; on many islands exploitation is traditional. Many birds die entangled in fishing nets; and there is some predation by feral cats (Felis catus) and rats (Rattus spp.). Mikura-jima islanders report that the population there is declining rapidly, probably due to predation by both rats and cats, but quantitative data are lacking (BirdLife International 2016), while potential predators present on some Korean nesting islands include rats (both Rattus rattus and R. norvegicus), cats, dogs and Siberian Weasals Mustela sibirica (55). Oil spills may also affect the species in Korean waters, although evidence for this is currently lacking (55). Significantly reduced levels of vitamin A have been found in nestlings on Mikura-jima, which has been linked to the fact that throughout incubation, and possibly also during the period when nestlings are fed, the majority of the global population forages in a marine area affected by fallout from the 2011 Fukushima nuclear plume (56). Protective measures are required, e.g. effective legal protection of species, combined with programmes of environmental education; fishing activities could be made less harmful to birds with suitable regulations concerning areas, seasons, methods and gear. Cats are being neutered and released on Mikura-jima (BirdLife International 2016). Not considered of conservation concern until 2015, when fears over possible population declines led to its listing as Near Threatened.
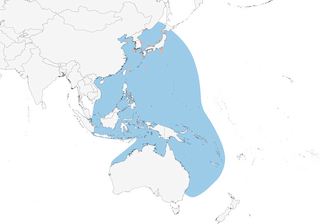
- Year-round
- Migration
- Breeding
- Non-Breeding


















