










Belford's Melidectes Melidectes belfordi Scientific name definitions
Text last updated January 1, 2008
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Catalan | menjamel de Belford |
| Dutch | Belfords Honingeter |
| English | Belford's Melidectes |
| English (United States) | Belford's Melidectes |
| Finnish | sininokimesikko |
| French | Méliphage de Belford |
| French (France) | Méliphage de Belford |
| German | Belfordhonigfresser |
| Indonesian | Melidektes belford |
| Japanese | アオガオヤマミツスイ |
| Norwegian | blåørehonningeter |
| Polish | dziwook modrolicy |
| Russian | Синелицый медосос |
| Serbian | Belfordov melidektes medojed |
| Slovak | medárik dlhozobý |
| Spanish | Mielero de Belford |
| Spanish (Spain) | Mielero de Belford |
| Swedish | belfordhonungsfågel |
| Turkish | Belford Balkuşu |
| Ukrainian | Медвянець синьощокий |
Melidectes belfordi (De Vis, 1890)
Definitions
- MELIDECTES
- belfordi
The Key to Scientific Names
Legend Overview
Originally Appeared in

Field Identification
26–29 cm; male 59–73 g and female 51–56 g (kinneari), male 70–80 g and female 55–67 g (other races). Nominate race has top and side of head black-brown, merging into light brownish-grey and faintly mottled dark brown on hindneck and side of neck, with short white supercilium bordering large pale blue orbital patch, latter broad behind eye and tapering to a point that joins a fine line of white feathers over rear ear-coverts; lower ear-coverts tinged grey and merge into brownish-grey side of neck; broad white malar stripe of elongated feathers from base of bill and broadening on side of upper throat; small white gape wattle; chin and upper throat black-brown, merging into brownish-grey or pale grey with brownish mottling on lower throat; mantle, back and scapulars black-brown, heavily scaled or mottled grey, merging into dark brown on rump and uppertail-coverts; upperwing and tail black-brown, yellow-olive tips and outer edges on greater coverts, broad yellow-olive outer edges on all except outer two remiges (large yellow-olive panel on folded wing), outer 5–6 primaries also narrowly tipped yellow-brown (reduced with wear), broad yellow-olive outer edges on rectrices (folded tail appears yellow-olive with dark centre); underbody pale grey, mottled dark brown and finely streaked white on breast and belly, merging to broader white barring or mottling on lower belly and vent, which also tinged rufous-brown, and to rufous-brown on undertail-coverts; undertail dark brown; underwing dark brown, slightly paler bases of remiges; iris brown to dark red-brown; bill black to dark greyish-black, sometimes dark-grey lower mandible; legs dark grey to dull blue-grey, varyingly mottled with brown or brownish-grey, soles yellow. Sexes alike in plumage, male larger than female. Juvenile is like adult but upperbody darker, sooty black-brown, with only faintly paler tips (mottled or scaled effect much reduced), underbody generally darker, blackish-grey, with much-reduced mottling or scaling throughout and a rufous tinge on belly, vent and undertail-coverts, also white malar stripe smaller, less white in line behind eye above rear ear-coverts. Races differ mainly in size and in plumage tones: brassi differs from nominate only in smaller size; kinneari is smaller than nominate, also paler grey-brown upperparts with paler and olive-tinged scaling on mantle, back and scapulars, brighter more yellowish-green edges of remiges, paler grey underbody with less prominent white markings posteriorly, and bare orbital skin violet-blue, also juvenile bare parts differ from adult’s, orbital skin greenish, gape yellow and legs brown (not brownish-grey to dark grey or blackish-brown); schraderensis differs from all others in having lemon-yellow (not blue) orbital ring, otherwise like previous in general size and coloration; joiceyi is smaller than others, and with strong olive tinge above.
Systematics History
Editor's Note: This article requires further editing work to merge existing content into the appropriate Subspecies sections. Please bear with us while this update takes place.
Sometimes treated as conspecific with M. leucostephes and with M. rufocrissalis. Co-occurs with latter on Mt Koimjim (Schrader Mts), but the two appear to exclude each other altitudinally; they hybridize freely, however, where they come into contact; Herzog Mts, much of Eastern Highlands, Mt Goliath (in E Star Mts) and N slopes of Snow Mts may once have supported both species, but hybrid populations now found extensively through these areas (although populations on N slopes of Snow Mts are almost pure individuals of present species, with only a few M. rufocrissalis characteristics). Race griseirostris, previously considered a hybrid form, now shown to be a stable population of hybrid origin (1), meriting recognition (2). Race brassi and nominate race may represent different species, as they occupy much the same geographical range but are separated altitudinally and appear not to intergrade; further study needed. Races joiceyi and kinneari intergrade in W Nassau Mts (Wissel Lakes area). Proposed race stresemanni (Herzog Mts) represents a hybrid form. Six subspecies recognized.Subspecies
Melidectes belfordi brassi Scientific name definitions
Distribution
Melidectes belfordi brassi Mayr & Rand, 1936
Definitions
- MELIDECTES
- belfordi
- brassi
The Key to Scientific Names
Legend Overview
Melidectes belfordi joiceyi Scientific name definitions

Distribution
Melidectes belfordi joiceyi (Rothschild, 1921)
Definitions
- MELIDECTES
- belfordi
- joiceyi
The Key to Scientific Names
Legend Overview
Melidectes belfordi kinneari Scientific name definitions
Distribution
Melidectes belfordi kinneari Mayr, 1936
Definitions
- MELIDECTES
- belfordi
- kinneari
The Key to Scientific Names
Legend Overview
Melidectes belfordi belfordi Scientific name definitions

Distribution
Melidectes belfordi belfordi (De Vis, 1890)
Definitions
- MELIDECTES
- belfordi
The Key to Scientific Names
Legend Overview
Melidectes belfordi schraderensis Scientific name definitions
Distribution
Melidectes belfordi schraderensis Gilliard & LeCroy, 1968
Definitions
- MELIDECTES
- belfordi
- schraderensis
The Key to Scientific Names
Legend Overview
Hybridization
Hybrid Records and Media Contributed to eBird
-
Belford's x Yellow-browed Melidectes (hybrid) Melidectes belfordi x rufocrissalis
Distribution
Editor's Note: Additional distribution information for this taxon can be found in the 'Subspecies' article above. In the future we will develop a range-wide distribution article.
Habitat
Primary upper montane to high mountain forests, forest edge, secondary growth and disturbed habitats, such as scattered trees in cattle pastures, patches of scrub in cleared areas and remnant forest; at high altitudes, recorded in thickets of shrubs in subalpine grassland. In upper Kaironk Valley, mainly in Nothofagus forest. Between Komopa and near Dauwagu, in W New Guinea, recorded also in Pandanus scrub and in open scrub with tree-ferns; at Dokfuma, in subalpine herbfield. Mainly above 1600 m, but recorded as low as 1400 m, and ranging up to tree-line in places, to 3800 m in SE, to at least 3350 m in CE and to above 3700 m in Snow Mts, probably up to 4000 m: in W of range, recorded at 3700–4000 m in Carstensz Mts and at 1740 m at Enarotali and 1830 m near Dauwagu; in E, not heard below 2300 m on Mt Kominjim (Schrader Mts) but common in higher forests to summit at c. 2600 m, and common on N side of upper Kaironk Valley from c. 2280 m upwards; on Mt Albert Edward from 2000 m to tree-line (at least 3350 m), but recorded down to 1600 m; in Victor Emanuel Mts at 2200–2300 m; in S foothills and slopes of Wharton Ranges, recorded at 1700–2700 m. Over much of range, replaced at lower altitudes by M. rufocrissalis (below 2300 m) or M. ochromelas.
Movement
No information; probably largely sedentary.
Diet and Foraging
Diet includes arthropods, mainly insects, including large ants (Formicidae) and beetles (Coleoptera); also nectar, fruit and, probably, pollen. Forages mainly in upper levels of trees, including emergents, and especially in flowering and fruiting trees (e.g. Banksia dentata, Xanthomyrtus papuanus) or flowering epiphytes (e.g. Rhododendron, Dimorphanthera); occasionally at ground level, and seen to forage at cluster (4-m diameter) of flowering Mitrastemma yamamotoi (a terrestrial root parasite), which birds visited mainly in mornings, usually descending cautiously and probing flowers while gripping vertical stem. Actions rather deliberate. Searches for insects by hopping along branches and up trunks, often head down, probing moss and epiphytes, and gleaning from foliage, vines and, apparently, flowers; often hangs to reach flowers, which it probes systematically. Conspicuous and pugnacious (but race kinneari considered surprisingly inconspicuous). Usually seen singly, sometimes in twos (probably pairs). Appears to defend resources from conspecifics and other birds; does not normally congregate in numbers in flowering trees, but may defend parts of trees. Sometimes associates with Melipotes fumigatus. Once seen to dive-bomb two Crested Berrypeckers (Paramythia montium). When foraging on Mitrastemma yamamotoi on ground, present species was found to be the most aggressive of four avian species, and effectively dominated and controlled foraging at the patch in the morning.
Sounds and Vocal Behavior
Breeding
Apparently predominantly late wet season and dry season: nest-building early Apr, late Jun and late Jul, “breeding birds” May and Aug–Oct, nestlings Apr–May and dependent fledglings early May and late Jun. Nest a deep cup of moss and twigs, densely lined with soft brownish material from base of tree-fern fronds, placed (not known whether suspended or supported) 4·5–9 m above ground in fork of branch in tree, one nest near Dauwagu c. 35 m above ground. No information on clutch size; one reported nest contained one young; both parents observed to feed nestlings and both seen with fledglings. No other information.
Conservation Status
Not globally threatened. Restrictedrange species: present in Central Papuan Mountains EBA. Common to very common in many parts of range, e.g. upper Kaironk Valley, Mt Koimjim. At English Peaks, at 3650 m in Wharton Range, 22 individuals mist-netted in 17 days. Interbreeding with M. rufocrissalis over wide area in E New Guinea; suggested that some hybridization occurs naturally in undisturbed habitats in narrow altitudinal band where the two species overlap, but that large-scale hybridization is a result of extensive habitat disturbance caused initially by indigenous agriculture and possibly other human activities in more recent times. Such habitat disturbance brings the two species into contact over broader areas and permits extensive and accelerated hybridization to take place.
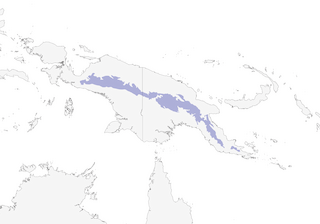
- Year-round
- Migration
- Breeding
- Non-Breeding

















