UPPERCASE: current genusUppercase first letter: generic synonym● and ● See: generic homonymslowercase: species and subspecies●: early names, variants, misspellings‡: extinct†: type speciesGr.: ancient GreekL.: Latin<: derived fromsyn: synonym of/: separates historical and modern geographic namesex: based onTL: type localityOD: original diagnosis (genus) or original description (species)
33–40 cm (1
Sun, Y.-H., Fang, Y., Swenson, J.E., Klaus, S. and Zheng, G.-M. (2005). Morphometrics of the Chinese grouse Bonasa sewerzowi. Journal of Ornithology. 146(1): 24–26.
); male 279–390 g, female 270–407 g (1
Sun, Y.-H., Fang, Y., Swenson, J.E., Klaus, S. and Zheng, G.-M. (2005). Morphometrics of the Chinese grouse Bonasa sewerzowi. Journal of Ornithology. 146(1): 24–26.
). Male
mass varies from lowest during breeding season to highest in winter, whereas the reverse is true in females (2
Sun, Y.-H. and Fang, Y. (2010). Chinese Grouse (Bonasa sewerzowi): its natural history, behavior and conservation. Chinese Birds. 1(3): 215–220.
). The world’s smallest grouse (1
Sun, Y.-H., Fang, Y., Swenson, J.E., Klaus, S. and Zheng, G.-M. (2005). Morphometrics of the Chinese grouse Bonasa sewerzowi. Journal of Ornithology. 146(1): 24–26.
). Very similar to B. bonasia but has less feathered tarsus (3
Madge, S., and P. McGowan (2002). Pheasants, Partridges and Grouse, including Buttonquails, Sandgrouse and Allies. Christopher Helm, London, UK.
); upperparts
more broadly barred black and upper breast
more chestnut, with only small white tips to feathers; tail has quite different barred pattern. Female
has less obvious red comb and further differs from male in being overall duller, with black throat largely obscured by whitish fringes to feathers
, and buffier underparts with less intense black barring (3
Madge, S., and P. McGowan (2002). Pheasants, Partridges and Grouse, including Buttonquails, Sandgrouse and Allies. Christopher Helm, London, UK.
). Irides dark brown, bill black, and feet greyish black (3
Madge, S., and P. McGowan (2002). Pheasants, Partridges and Grouse, including Buttonquails, Sandgrouse and Allies. Christopher Helm, London, UK.
). Juvenile plumage is apparently unknown (3
Madge, S., and P. McGowan (2002). Pheasants, Partridges and Grouse, including Buttonquails, Sandgrouse and Allies. Christopher Helm, London, UK.
). Race secunda is described as being more rufous than nominate.
Systematics History
Editor's Note: This article requires further editing work to merge existing content into the appropriate Subspecies sections. Please bear with us while this update takes place.
Formerly placed in genus Tetrastes, but genetic evidence clearly places both present species and closely related B. bonasia in Bonasa together with North American B. umbellus (4
Lucchini, V., Höglund, J., Klaus, S., Swenson, J. and Randi, E. (2001). Historical biogeography and a mitochondrial DNA phylogeny of grouse and ptarmigan. Mol. Phyl. & Evol.. 20(1): 149–162.
), despite some behavioural differences between the American and the two Eurasian species (5
Drovetski, S.V., Rohwer, S. and Mode, N.A. (2006). Role of sexual and natural selection in evolution of body size and shape: a phylogenetic study of morphological radiation in grouse. J. Evol. Biol. 19(4): 1083–1091.
). Geographical variation slight. S race secunda slightly more rufous, but this is perhaps no more than clinal variation (3
Madge, S., and P. McGowan (2002). Pheasants, Partridges and Grouse, including Buttonquails, Sandgrouse and Allies. Christopher Helm, London, UK.
). Two subspecies tentatively recognized.
Subspecies
Monotypic.
Distribution
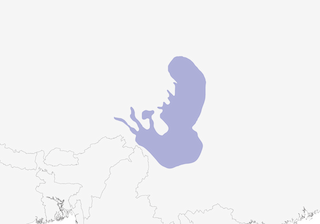
Mountains of C China, from C Gansu and S Qinghai to E Tibet, NW Yunnan and N Sichuan.
Habitat
Birch and coniferous mountain forests, generally above 2400 m, and as high as 4300 m in Tibet (6
Sun, Y-H. (2000). Distribution and status of the Chinese grouse Bonasa sewerzowi. Wildl. Biol.. 6: 271–275.
); the elevation occupied increases with decreasing latitude, e.g. breeds at 2350–3000 m in Qilian Mts of NW Gansu, but at 3530–3820 m in NW Yunnan. Records as low as 1000 m are probably doubtful (6
Sun, Y-H. (2000). Distribution and status of the Chinese grouse Bonasa sewerzowi. Wildl. Biol.. 6: 271–275.
). In places, may reach treeline and during summer even alpine belt, in areas of rhododendron. In Gansu, main habitat types centred on dragon spruce (Picea crassifolia or P. asperata) and Abies fargesii (1
Sun, Y.-H., Fang, Y., Swenson, J.E., Klaus, S. and Zheng, G.-M. (2005). Morphometrics of the Chinese grouse Bonasa sewerzowi. Journal of Ornithology. 146(1): 24–26.
), sometimes with 60–90% cover of moss (Abietinella, Bryum) on forest floor; other typical plants include birch (Betula), willow (Salix) and barberry (Berberis); grassland and alpine shrubs generally avoided. Females with broods chose sites with significantly less canopy cover, greater willow cover and greater herb height, using a variety of vegetation types at different stages of chick development (7
Zhao, J.-M., Fang, Y., Lou, Y.-Q. and Sun, Y.-H. (2015). Brood habitat selection of Chinese Grouse (Tetrastes sewerzowi) at Lianhuashan, Gansu, China. Wilson J. Orn.. 127(2): 310–318.
).
Movement
Apparently sedentary. Mean territory size of paired male in spring is c. 2·1 ha (range in one study 1·8–3·2 ha) (8
Sun, Y.-H. and Fang, Y. (1997). Notes on the natural history and behaviour of the Chinese grouse Bonasa sewerzowi. Wildl. Biol.. 3: 265–268.
), and of unpaired male c. 2·7 ha (9
Lu, N. and Sun, Y.-H. (2011). Population viability analysis and conservation of Chinese Grouse Bonasa sewerzowi in Lianhuashan Nature Reserve, north-west China. Bird Conservation International. 21(1): 49–58.
). Radio-tracking data reveal that home ranges of males in winter range from 6 ± 1·6 ha (n = 9) to 16 ± 2·6 ha (n = 10); most females and all yearlings form mixed-sex flocks at this season (between mid Oct and mid Mar) (3
Madge, S., and P. McGowan (2002). Pheasants, Partridges and Grouse, including Buttonquails, Sandgrouse and Allies. Christopher Helm, London, UK.
), whereas adult males either remain on breeding territory, move to winter flocking area (where groups typically number 4–5 to 13–14 individuals) (10
Sun, Y.-H. (1997). Winter flocking behaviour of Chinese grouse Bonasa sewerzowi. Wildl. Biol.. 3(3–4): 290.
), or wander between spring territory and wintering area (10
Sun, Y.-H. (1997). Winter flocking behaviour of Chinese grouse Bonasa sewerzowi. Wildl. Biol.. 3(3–4): 290.
, 11
Yang, C., Fang, Y. and Sun, Y.-H. (2011). Winter space use and social behaviors of Chinese Grouse (Bonasa sewerzowi) at Lianhuashan mountains, Gansu, China. Journal of Ornithology. 152(2): 297–305.
), but females can also briefly form territories in autumn (12
Klaus, S., Sun, Y.-H., Fang, Y. and Scherzinger, W. (2009). Autumn territoriality of Chinese Grouse Bonasa sewerzowi at Lianhuashan Natural Reserve, Gansu, China. Int. Journal Galliform Conserv. 1: 44–48.
). Those males that join flocks in winter leave these earlier in spring (2
Sun, Y.-H. and Fang, Y. (2010). Chinese Grouse (Bonasa sewerzowi): its natural history, behavior and conservation. Chinese Birds. 1(3): 215–220.
).
Diet and Foraging
In Gansu, mainly flower and leaf buds of willow and birch in early spring, moving onto flower buds and leaves of the same species by May; in Jun–Jul, seeds and inflorescences of Polygonum dominate, while for brief period in autumn mountain ash (Sorbus) berries are clearly preferred (13
Klaus, S., Lou, Y., Fang, Y., Scherzinger, W. and Sun, Y.-H. (2014). Aggressive interactions between males of Chinese Grouse Tetrastes sewerzowi in autumn at Lianhuashan natural reserve, Gansu, central China. Orn. Anz.. 53(1–2): 45–53.
); between Jan and Mar, sea buckthorn (Hippophae rhamnoides) is important (10
Sun, Y.-H. (1997). Winter flocking behaviour of Chinese grouse Bonasa sewerzowi. Wildl. Biol.. 3(3–4): 290.
), but willow buds and twigs (83% of foraging observations) of six main species are still the most important constituent, of which Kangding willow (Salix paraplesia) is the most preferred species and Sichuan willow (S. hylonoma) the least (14
Wang, J., Fang, Y., Klaus, S. and Sun, Y.-H. (2012). Winter foraging strategy of the Chinese Grouse (Bonasa sewerzowi): ecological and physiological factors. Journal of Ornithology. 153(2): 257–264.
). In late Apr–May willows are the primary food of males (> 98% wet biomass), while spruce (Picea aspera) seeds and willow are primary food of females, which also eat frequently invertebrates (mainly ants) and forbs (possibly to meet the nutrient demands of egg formation) (15
Jie, W., Chen,Y., Nan, L., Yun, F. and Yue-Hua, S. (2010). Diet of Chinese Grouse (Tetrastes sewerzowi) during preincubation. Wilson J. Orn.. 122(1): 177–180.
). Forages on ground or in trees.
Sounds and Vocal Behavior
Territorial displays are given in both spring and autumn (2
Sun, Y.-H. and Fang, Y. (2010). Chinese Grouse (Bonasa sewerzowi): its natural history, behavior and conservation. Chinese Birds. 1(3): 215–220.
). Males perform a wingbeat display, with variable flutter-jumps (the wings making a “pu pu pu pu” sound) (3
Madge, S., and P. McGowan (2002). Pheasants, Partridges and Grouse, including Buttonquails, Sandgrouse and Allies. Christopher Helm, London, UK.
) from the ground reaching up to 1 m high and 2–4 m wide, sometimes also from a tree to the ground and vice versa, especially immediately after leaving the the roost until c. 08:00 hours (16
Klaus, S., Scherzinger, W. and Sun, Y.-H. (1998). Territorial- und Werbeverhalten des Chinahaselhuhns (Bonasa sewerzowi). Journal of Ornithology. 139(2): 185–186.
); this display serves to advertise territory and attract females, but perhaps also as a warning signal (8
Sun, Y.-H. and Fang, Y. (1997). Notes on the natural history and behaviour of the Chinese grouse Bonasa sewerzowi. Wildl. Biol.. 3: 265–268.
). Noisy territorial flights in the crowns of trees are rare (16
Klaus, S., Scherzinger, W. and Sun, Y.-H. (1998). Territorial- und Werbeverhalten des Chinahaselhuhns (Bonasa sewerzowi). Journal of Ornithology. 139(2): 185–186.
). The low-frequency sounds generated by the wings produce an impressive concert, especially if 3–4 conflicting males are in close proximity (16
Klaus, S., Scherzinger, W. and Sun, Y.-H. (1998). Territorial- und Werbeverhalten des Chinahaselhuhns (Bonasa sewerzowi). Journal of Ornithology. 139(2): 185–186.
). Male–male aggression displays include hoarse “en er en er en...” sounds comprising separate trill and squeak phases comprising three and 4–7 elements, respectively, sometimes with a cackling sound given after the trill (13
Klaus, S., Lou, Y., Fang, Y., Scherzinger, W. and Sun, Y.-H. (2014). Aggressive interactions between males of Chinese Grouse Tetrastes sewerzowi in autumn at Lianhuashan natural reserve, Gansu, central China. Orn. Anz.. 53(1–2): 45–53.
), while females utter a “ze ze ze-dackdack” in company of chicks to alert them to potential predators (3
Madge, S., and P. McGowan (2002). Pheasants, Partridges and Grouse, including Buttonquails, Sandgrouse and Allies. Christopher Helm, London, UK.
), but four different types of alarm calls have been identified (17
Shi, M., Sun, Y.-H., Wen, A.-X. and Fang, Y. (2013). Spring territory behavior of the Chinese Grouse (Tetrastes sewerzowi) at Lianhuashan, Gansu, China. Chinese J. Zool.. 48(5): 665–672. (In Chinese with English summary.).
), including a longer “tetetetete... ” uttered by a female during an encounter with males (13
Klaus, S., Lou, Y., Fang, Y., Scherzinger, W. and Sun, Y.-H. (2014). Aggressive interactions between males of Chinese Grouse Tetrastes sewerzowi in autumn at Lianhuashan natural reserve, Gansu, central China. Orn. Anz.. 53(1–2): 45–53.
).
.
Breeding
Laying from end May to mid Jun (3
Madge, S., and P. McGowan (2002). Pheasants, Partridges and Grouse, including Buttonquails, Sandgrouse and Allies. Christopher Helm, London, UK.
). Monogamous; although polyandry is occasionally suspected (8
Sun, Y.-H. and Fang, Y. (1997). Notes on the natural history and behaviour of the Chinese grouse Bonasa sewerzowi. Wildl. Biol.. 3: 265–268.
, 18
Sun, Y.-H. and Fang, Y. (1998). The mating system of Chinese Grouse Bonasa sewerzowi. P. 309 in:.
), only available evidence suggests strong pair-bonds (2
Sun, Y.-H. and Fang, Y. (2010). Chinese Grouse (Bonasa sewerzowi): its natural history, behavior and conservation. Chinese Birds. 1(3): 215–220.
); male territories established in late winter (3
Madge, S., and P. McGowan (2002). Pheasants, Partridges and Grouse, including Buttonquails, Sandgrouse and Allies. Christopher Helm, London, UK.
), but pairs form in late Mar (18
Sun, Y.-H. and Fang, Y. (1998). The mating system of Chinese Grouse Bonasa sewerzowi. P. 309 in:.
), with copulation observed from second week of May (16
Klaus, S., Scherzinger, W. and Sun, Y.-H. (1998). Territorial- und Werbeverhalten des Chinahaselhuhns (Bonasa sewerzowi). Journal of Ornithology. 139(2): 185–186.
). As many as 44% of males did not mate in one study (19
Sun, Y.-H., Swenson, J.E., Fang, Y., Klaus, S. and Scherzinger, W. (2003). Population ecology of the Chinese grouse, Bonasa sewerzowi, in a fragmented landscape. Biological Conservation. 110: 177–184.
). Nest scrape (c. 43 mm by 30·5 mm) (3
Madge, S., and P. McGowan (2002). Pheasants, Partridges and Grouse, including Buttonquails, Sandgrouse and Allies. Christopher Helm, London, UK.
) at base of tree, in tree stump or on rocky ledge; constructed of grass, sometimes lined with pine needles and partly edged with moss; of 103 nests found in one study, 56% were at base of deciduous trees, mainly willows (Salix spp.) and birches (Betula utilis), and 40% at base of conifers, typically sited on N- or E-facing slopes and 88% of nests were on slopes < 30º, while females often nested within c. 150 m of the previous year’s site (20
Sun, Y.-H., Fang, Y., Jia, C-X., Klaus, S., Swenson, J.E. and Scherzinger, W. (2007). Nest site selection of Chinese grouse Bonasa sewerzowi at Lianhuashan, Gansu, China. Wildl. Biol.. 13(Suppl. 1): 68–72.
). Will occasionally re-nest if first clutch is lost (19
Sun, Y.-H., Swenson, J.E., Fang, Y., Klaus, S. and Scherzinger, W. (2003). Population ecology of the Chinese grouse, Bonasa sewerzowi, in a fragmented landscape. Biological Conservation. 110: 177–184.
). Clutch 5–8 (mean 6·19 in one study) (1
Sun, Y.-H., Fang, Y., Swenson, J.E., Klaus, S. and Zheng, G.-M. (2005). Morphometrics of the Chinese grouse Bonasa sewerzowi. Journal of Ornithology. 146(1): 24–26.
) pale yellow (1
Sun, Y.-H., Fang, Y., Swenson, J.E., Klaus, S. and Zheng, G.-M. (2005). Morphometrics of the Chinese grouse Bonasa sewerzowi. Journal of Ornithology. 146(1): 24–26.
) or yellowish-brown (3
Madge, S., and P. McGowan (2002). Pheasants, Partridges and Grouse, including Buttonquails, Sandgrouse and Allies. Christopher Helm, London, UK.
) eggs with diffuse chestnut markings (3
Madge, S., and P. McGowan (2002). Pheasants, Partridges and Grouse, including Buttonquails, Sandgrouse and Allies. Christopher Helm, London, UK.
), size 40–43·5 mm × 30·8–31·5 mm (1
Sun, Y.-H., Fang, Y., Swenson, J.E., Klaus, S. and Zheng, G.-M. (2005). Morphometrics of the Chinese grouse Bonasa sewerzowi. Journal of Ornithology. 146(1): 24–26.
), mean mass 20·5 g, laid every 48 hours (2
Sun, Y.-H. and Fang, Y. (2010). Chinese Grouse (Bonasa sewerzowi): its natural history, behavior and conservation. Chinese Birds. 1(3): 215–220.
); incubation 25 days (in captivity) by female alone (3
Madge, S., and P. McGowan (2002). Pheasants, Partridges and Grouse, including Buttonquails, Sandgrouse and Allies. Christopher Helm, London, UK.
), but 27–29 days in wild (2
Sun, Y.-H. and Fang, Y. (2010). Chinese Grouse (Bonasa sewerzowi): its natural history, behavior and conservation. Chinese Birds. 1(3): 215–220.
); downy chicks have head markings more apparent than in other Bonasa, tended by female alone, remain with adult for 2–3 months and then disperse up to 5 km (2
Sun, Y.-H. and Fang, Y. (2010). Chinese Grouse (Bonasa sewerzowi): its natural history, behavior and conservation. Chinese Birds. 1(3): 215–220.
); mean 3·62 young/year per female fledged in one study (19
Sun, Y.-H., Swenson, J.E., Fang, Y., Klaus, S. and Scherzinger, W. (2003). Population ecology of the Chinese grouse, Bonasa sewerzowi, in a fragmented landscape. Biological Conservation. 110: 177–184.
); nest loss is 37%, of which 10–29% is caused by egg collecting by local people (19
Sun, Y.-H., Swenson, J.E., Fang, Y., Klaus, S. and Scherzinger, W. (2003). Population ecology of the Chinese grouse, Bonasa sewerzowi, in a fragmented landscape. Biological Conservation. 110: 177–184.
), but hatching success in one study was 93% (2
Sun, Y.-H. and Fang, Y. (2010). Chinese Grouse (Bonasa sewerzowi): its natural history, behavior and conservation. Chinese Birds. 1(3): 215–220.
). Mortality suspected to be c. 68% within first year of life, declining to 34–39% per annum thereafter (9
Lu, N. and Sun, Y.-H. (2011). Population viability analysis and conservation of Chinese Grouse Bonasa sewerzowi in Lianhuashan Nature Reserve, north-west China. Bird Conservation International. 21(1): 49–58.
), although one study reported that only 17% of chicks survive to 13 weeks old (19
Sun, Y.-H., Swenson, J.E., Fang, Y., Klaus, S. and Scherzinger, W. (2003). Population ecology of the Chinese grouse, Bonasa sewerzowi, in a fragmented landscape. Biological Conservation. 110: 177–184.
). Potential mammalian predators of eggs and/or adults include dhole (Cuon alpinus), raccoon dog (Nycterentes procyonoides), stone marten (Martes foina), Siberian weasel (M. sibirica), sand badger (Arctomyx collaris), Pallas’ cat (Felis manul), leopard cat (F. bengalensis) and Eurasian lynx (Lynx lynx), while among birds various corvids, owls and diurnal raptors inflict even more severe losses among adults (19
Sun, Y.-H., Swenson, J.E., Fang, Y., Klaus, S. and Scherzinger, W. (2003). Population ecology of the Chinese grouse, Bonasa sewerzowi, in a fragmented landscape. Biological Conservation. 110: 177–184.
). Males thought to achieve sexual maturity when two years old, but females a year earlier (9
Lu, N. and Sun, Y.-H. (2011). Population viability analysis and conservation of Chinese Grouse Bonasa sewerzowi in Lianhuashan Nature Reserve, north-west China. Bird Conservation International. 21(1): 49–58.
). Overall population is thought to be skewed towards males, 64% versus 36% in females, at least locally (9
Lu, N. and Sun, Y.-H. (2011). Population viability analysis and conservation of Chinese Grouse Bonasa sewerzowi in Lianhuashan Nature Reserve, north-west China. Bird Conservation International. 21(1): 49–58.
). Maximum recorded longevity just over five years (19
Sun, Y.-H., Swenson, J.E., Fang, Y., Klaus, S. and Scherzinger, W. (2003). Population ecology of the Chinese grouse, Bonasa sewerzowi, in a fragmented landscape. Biological Conservation. 110: 177–184.
).
Not globally threatened. Currently considered Near Threatened. Listed as endangered in Chinese Red Data book. Total range speculated to be c. 394,000 km² and population c. 10,000 birds (21
Storch, I. (2007). Grouse: Status Survey and Conservation Action Plan 27747746–27741774. IUCN, Gland & World Pheasant Association, Fordingbridge.
); range recently (1997) (6
Sun, Y-H. (2000). Distribution and status of the Chinese grouse Bonasa sewerzowi. Wildl. Biol.. 6: 271–275.
) discovered to be more extensive than previously thought in W, and species may yet prove to be widespread in suitable habitat in SE Tibet. Very few data available, although knowledge of the species has noticeably advanced in recent decades. Known from at least 18 protected areas across range, of which eight are in Sichuan, six Gansu and two each in Qinghai and Yunnan (6
Sun, Y-H. (2000). Distribution and status of the Chinese grouse Bonasa sewerzowi. Wildl. Biol.. 6: 271–275.
). Common in suitable forest, e.g. density of 15 occupied territories/km² in Lianhuashan Natural Reserve (S Gansu) (16
Klaus, S., Scherzinger, W. and Sun, Y.-H. (1998). Territorial- und Werbeverhalten des Chinahaselhuhns (Bonasa sewerzowi). Journal of Ornithology. 139(2): 185–186.
) and 4–6 pairs/ha at Zhongdian (Yunnan) but just 0·27–2 birds/km² at Baihe Natural Reserve (Sichuan) (6
Sun, Y-H. (2000). Distribution and status of the Chinese grouse Bonasa sewerzowi. Wildl. Biol.. 6: 271–275.
); species has disappeared from E Qinghai and C Gansu, due to deforestation, and is also experiencing intensive habitat loss in SW Gansu; climate change, which is speculated to lead to increasing aridity within the species’ range, is also cause for concern (22
Klaus, S., Scherzinger, W. and Sun, Y.-H. (1996). Ökologie und Verhalten des Chinahaselhuhns Bonasa sewerzowi. Orn. Beob.. 93: 343–365.
). Can probably persist in selectively logged forest, albeit at obviously lower densities (6
Sun, Y-H. (2000). Distribution and status of the Chinese grouse Bonasa sewerzowi. Wildl. Biol.. 6: 271–275.
), with habitat fragmentation (as well as degradation and outright loss) an additional risk (2
Sun, Y.-H. and Fang, Y. (2010). Chinese Grouse (Bonasa sewerzowi): its natural history, behavior and conservation. Chinese Birds. 1(3): 215–220.
, 23
Lu, N., Jia, C.-X., Lloyd, H. and Sun, Y.-H. (2012). Species-specific habitat fragmentation assessment, considering the ecological niche requirements and dispersal capability. Biological Conservation. 152: 102–109.
), as species prefers patches larger than 40 ha and suitable but unoccupied habitat fragments isolated by > 2 km are apparently inaccessible to the species (24
Klaus, S., Voigt, W., Fang, Y., Selsam, P. and Sun, Y.-H. (2013). Patch occupancy by the Chinese Grouse (Tetrastes sewerzowi) in a fragmented landscape. Chinese Birds. 4(3): 232–239.
). Significant predation of eggs by Large-billed Crows (Corvus macrorhynchos), accounting for failure of 50% of nests located during recent study, but egg-collecting by local people, even within protected areas is also an important threat (9
Lu, N. and Sun, Y.-H. (2011). Population viability analysis and conservation of Chinese Grouse Bonasa sewerzowi in Lianhuashan Nature Reserve, north-west China. Bird Conservation International. 21(1): 49–58.
). Illegal hunting also occurs (9
Lu, N. and Sun, Y.-H. (2011). Population viability analysis and conservation of Chinese Grouse Bonasa sewerzowi in Lianhuashan Nature Reserve, north-west China. Bird Conservation International. 21(1): 49–58.
), with some people admitting to shooting 60–80 birds per year (21
Storch, I. (2007). Grouse: Status Survey and Conservation Action Plan 27747746–27741774. IUCN, Gland & World Pheasant Association, Fordingbridge.
).
de Juana, E. and G. M. Kirwan (2020). Severtzov's Grouse (Tetrastes sewerzowi), version 1.0. In Birds of the World (J. del Hoyo, A. Elliott, J. Sargatal, D. A. Christie, and E. de Juana, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.sevgro1.01
Partnerships
A global alliance of nature organizations working to document the natural history of all bird species at an unprecedented scale.