










Sardinian Warbler Curruca melanocephala Scientific name definitions
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Albanian | Bilbilthi kokëzi gushëbardhë |
| Arabic | هازجة سردينيا الرأساء |
| Asturian | Papuda tiestaprieta |
| Basque | Txinbo burubeltza |
| Bulgarian | Малко черноглаво коприварче |
| Catalan | tallarol capnegre |
| Croatian | crnoglava grmuša |
| Czech | pěnice bělohrdlá |
| Danish | Sorthovedet Sanger |
| Dutch | Kleine Zwartkop |
| English | Sardinian Warbler |
| English (United States) | Sardinian Warbler |
| Finnish | samettipääkerttu |
| French | Fauvette mélanocéphale |
| French (France) | Fauvette mélanocéphale |
| Galician | Papuxa de cabeza negra |
| German | Samtkopf-Grasmücke |
| Greek | Μαυροτσιροβάκος |
| Hebrew | סבכי שחור-ראש |
| Hungarian | Kucsmás poszáta |
| Icelandic | Hjálmsöngvari |
| Italian | Occhiocotto |
| Japanese | クロガシラムシクイ |
| Lithuanian | Juodagalvė sardininė devynbalsė |
| Norwegian | svarthodesanger |
| Polish | pokrzewka aksamitna |
| Portuguese (Portugal) | Toutinegra-dos-valados |
| Romanian | Silvie mediteraneană |
| Russian | Средиземноморская славка |
| Serbian | Sredozemna crnoglava grmuša |
| Slovak | penica sivá |
| Slovenian | Žametna penica |
| Spanish | Curruca Cabecinegra |
| Spanish (Spain) | Curruca cabecinegra |
| Swedish | sammetshätta |
| Turkish | Maskeli Ötleğen |
| Ukrainian | Кропив’янка середземноморська |
Curruca melanocephala (Gmelin, 1789)
Definitions
- CURRUCA
- curruca
- melanocephala / melanocephalon / melanocephalos / melanocephalum / melanocephalus
The Key to Scientific Names
Legend Overview
Field Identification
13·5 cm; 12–25 g. Male nominate race breeding has glossy black head contrasting with white chin and throat; upperparts and upperwing-coverts dark ash-grey, mantle and scapulars tinged brown; remiges blackish with grey fringes; tail blackish with white to grey fringes and white tips and edges; underparts greyish-white; iris red-brown and orbital ring reddish , both contrasting with pinkish-orange eyering; bill blackish, bluish base of lower mandible; legs brownish. Male in non-breeding plumage is similar to breeding, but upperparts more olive and head less blackish, especially at rear. Female breeding has medium-grey to dark grey head well demarcated from dull white chin and throat, upperparts variably brownish , tail as male but white areas duller and with less contrasting pattern, underparts cream-buff with olive-brown sides, iris, orbital ring and eyering normally somewhat paler than male; female non-breeding similar to breeding, but head more profusely tinged brownish and thus less well demarcated from upperparts. Juvenile is similar to female but duller and less contrasted, head brownish (concolourous with rest of upperparts in female but greyer in male), upperparts brown, wing browner than adult and fringed buffish, throat and underparts extensively buffish, iris greyish, orbital ring pale reddish; first-winter like respective adult, but with juvenile-like bare parts and unmoulted flight-feathers. Racial differences rather marked, involving size and plumage coloration: <em>leucogastra</em> is smaller than nominate, has more rounded wings than other races and less white on inner webs of outer rectrices, and overall paler on E islands (Lanzarote, Fuerteventura, Gran Canaria) and darker on those of W (Palma, Tenerife ); momus is smaller and paler overall; valverdei is similar in size to nominate, but has palest upperparts and lacks dark grey tones or brown suffusion, adult male with distinctly matt black (not glossy) cap.
Systematics History
Editor's Note: This article requires further editing work to merge existing content into the appropriate Subspecies sections. Please bear with us while this update takes place.
Geographical variation within each race largely clinal, becoming more complex in insular areas (especially Canary Is); race leucogastra sometimes synonymized with nominate, but detailed study (1) suggests its revalidation, although this rejected by others (2, 3); variation (in colour and size) within islands, however, noticeable and demands further work. Recently described race valverdei (1) represents a new stage in variability of the species, probably related to particular ecological conditions in Sahara Desert. Proposed races pasiphae (described from Crete) and carmichaellowi (SE Italy) considered to fall within range of variation of nominate, and included within it. Race †norrisae extinct. Four extant subspecies recognized.Subspecies
Curruca melanocephala melanocephala Scientific name definitions
Distribution
Curruca melanocephala melanocephala (Gmelin, 1789)
Definitions
- CURRUCA
- curruca
- melanocephala / melanocephalon / melanocephalos / melanocephalum / melanocephalus
The Key to Scientific Names
Legend Overview
Curruca melanocephala norrisae Scientific name definitions
Distribution
Curruca melanocephala norrisae (Nicoll, 1917)
Definitions
- CURRUCA
- curruca
- melanocephala / melanocephalon / melanocephalos / melanocephalum / melanocephalus
- norrisae
The Key to Scientific Names
Legend Overview
Curruca melanocephala momus Scientific name definitions

Distribution
Curruca melanocephala momus Hemprich & Ehrenberg, 1833
Definitions
- CURRUCA
- curruca
- melanocephala / melanocephalon / melanocephalos / melanocephalum / melanocephalus
- momus
- Momus
The Key to Scientific Names
Legend Overview
Distribution
Editor's Note: Additional distribution information for this taxon can be found in the 'Subspecies' article above. In the future we will develop a range-wide distribution article.
Habitat
Probably one of the most generalist of Mediterranean warblers: present in all types of warm habitat at low altitudes, and up to c. 1200–1300 m in most of range and to 1800 m in NW Africa. Occupies maquis, garrigue, dry coastal habitats, urban and suburban environments (including gardens ), abandoned farmland and dry coast sites; present also in Mediterranean cultures such as olives, almonds, orchards, citrus groves, vineyards, but favours mosaic landscapes with hedgerows, bushes and stands of pines, mostly Aleppo pine (Pinus halepensis); common in holm oak (Quercus ilex) forests, especially low dense forest with bushy undergrowth. Despite apparently unfavourable conditions can recolonize burnt Mediterranean habitats, sometimes as early as second year after wildfire. Race valverdei said to occur in three distinct desert habitats in Western Sahara: Euphorbia and Opuntia inermis vegetation; stands of tamarisk (Tamarix) and Nitrata alternating with Salicornia; and “graras”, defined as concentric circles of vegetation formed by Euphorbia, saltworts (Salsola), Rhus and, in centre, grass. During winter in non-breeding areas of Morocco, Mauritania, Senegal and Sudan, occurs in desert shrubs, gardens, tamarisks, edges of mangrove and dry acacia (Acacia) grassland; the second most abundant passerine in some oases of Algeria and also seen very often in acacia trees and Zilla spinosa bush; in Tunisia reported as very abundant in N & C of the country, with highest densities in wild olive and Lentiscus vegetation and in oak and thuya (Tetraclinis articulata) maquis.
Movement
Partially migratory. Island and coastal populations mostly sedentary, but proportion of migrants increases inland and to N and, especially, E (where N populations appear to be entirely migratory). As migrants and residents overlap in much of range, difficult to distinguish phenology of migrants; nevertheless, increase in numbers during autumn and spring and presence of individuals in non-breeding areas allow detection of passage. Majority of movements short, but long-distance migration certainly occurs, as demonstrated by presence of wintering birds S of Sahara; non-breeding range extends S in Africa to N Senegal in W and to N Sudan in E (in discontinuous zone). Post-breeding migration late Aug to Dec, with peaks in W Europe mostly during Oct–Nov; in Turkey some areas completely vacated by late Aug, and in Cyprus first migrants recorded from late Oct onwards; in Israel migration mainly Nov–Dec; reaches wintering grounds in Mauritania and Niger in Oct. Ringing recoveries of nominate race include movements between Balearics and Iberian Peninsula and even between continental Europe and NW Africa. Return to breeding areas takes place from late Feb to Apr; some variation between years and populations involved; migration along Morocco and Algeria coast recorded mostly in Mar. During autumn, local movements probably common in all populations as a result of juvenile dispersal and variations in availability of seasonal food resources, such as berries. In Israel, immatures of local breeding race (momus) recorded as performing altitudinal movements.
Diet and Foraging
Diet mainly arthropods ; high amount of fruit, especially in autumn and winter , but also throughout year, probably related to local availability. Invertebrate food includes insects and their larvae from a wide array of orders, also spiders (Araneae) and sometimes also small snails; beetles (Coleoptera) and hymenopterans recorded in large percentages in some studies. In study in S France, 97% of stomachs contained invertebrates , including 70% beetles, 27% ants and 18% snails. On S wintering grounds in Niger, earwigs (Dermaptera), termites (Isoptera), caterpillers, hymenopterans (wasps, ants) and beetles recorded in diet. Among fruits, those of Rhamnus, Ficus, Rubus, Olea and Pistacea consumed throughout range. In Spain, fruits of the buckthorn Rhamnus alaeternus consumed regularly by adults during breeding season and also fed to nestlings. Olives an important source of food during winter; small olives swallowed whole, larger ones partially pecked. In Israel, c. 84% of annual crop of honeysuckle (Lonicera) fruits consumed by this species. Nectar consumption also frequent in late winter and early spring and probably more regular than was previously thought; individuals sometimes defend groups of plants against smaller birds. Recorded also as eating bread and peanuts , and in winter occasionally visits bird-feeders. Forages mostly in scrub, also on ground and in low trees.
Sounds and Vocal Behavior
Song , from top of bush or in display-flight, a musical and varied rapid chatter 2–5 seconds in duration, typically involves “t-r”-like notes with short whistles; repertoire rich with the improvisation of new notes once the song is progressing. Most usual call a distinctive “t-tra, t-tra, t-tra” , frequently accompanied by loud rattling “tr-tr-tr”.
Breeding
Season chiefly Mar–Jun, some pairs earlier, in Feb, and one report from Morocco of adult still feeding fledged young in late Oct; 1–3 broods, mostly two. Monogamous; solitary, territorial breeder. Resident male starts song flights as early as Feb, rising 2–3 m above ground and gliding, with wings raised high, to same or another perch. Nest built mainly by female, a grass cup placed usually 30–60 cm above ground (exceptionally, up to 3 m high) in scrub or bush or, less frequently, among dense herbs or in small tree; in Spain most nests at low height in kermes oak (Quercus coccifera), in Morocco often almost at ground level in Inula viscosa or Chamaerops humilis; mean territory size estimated at 8779 m². Clutch 3–5 eggs , early and late clutches smaller; incubation by both sexes, period 12–15 days; chicks fed by both parents, nestling period 12–13 days; fledglings remain with parents for 2–3 weeks. In Malta breeding success estimated at 44%. Oldest recorded individual 7 years 7 months; annual adult survival at least 55% in Gibraltar.
Conservation Status
Not globally threatened. Locally common to very common; in favourable habitats sometimes the most abundant avian species. Estimated European population of over 5,800,000 breeding pairs seems clearly lower than the true figure, as judged from local abundance and more precise regional estimates. Densities of up to 7·4–10 breeding pairs/10 ha. Race norrisae was restricted to Faiyum (in N Egypt), where was common until 1920s, but last recorded in 1939 and now considered extinct; demise apparently due to habitat loss and salination. Has expanded its breeding distribution since 19th century in some countries, i.e. Spain, France, Italy, Bulgaria, Romania, Malta and Cyprus. Extremely harsh winters can affect this species’ survival (mainly in N parts of range); thus, winters of 1946–47 and 1962–63 had adverse impact on population in S France, and 2001–02 winter caused descrease in NE Spain, in some cases with declines in breeding numbers of up to 50%. Has been suggested that current global climatic warming could benefit this species in Europe. Conversely, global warming has been suggested as cause of decline in body mass and body condition observed for this and other sedentary passerines in Israel between 1950 and 1999.
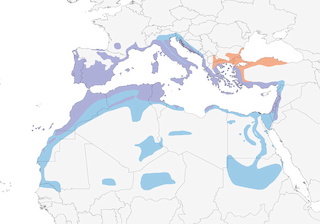
- Year-round
- Migration
- Breeding
- Non-Breeding


eBird and Macaulay Library

eBird Status and Trends

















