










White-breasted Waterhen Amaurornis phoenicurus Scientific name definitions
Revision Notes
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Afrikaans | Witborsriethaan |
| Assamese | ডাউক |
| Bangla | ডাহুক |
| Catalan | polla pitblanca |
| Chinese | 白腹秧雞 |
| Chinese (Hong Kong SAR China) | 白胸苦惡鳥 |
| Chinese (SIM) | 白胸苦恶鸟 |
| Czech | chřástal běloprsý |
| Dutch | Witborstwaterhoen |
| English | White-breasted Waterhen |
| English (United States) | White-breasted Waterhen |
| French | Râle à poitrine blanche |
| French (France) | Râle à poitrine blanche |
| German | Weißbrust-Kielralle |
| Gujarati | સફેદછાતી સંતાકૂકડી |
| Hebrew | סופית לבנת-חזה |
| Hindi | सफ़ेद-छाती जलमुर्गी |
| Hungarian | Fehérmellű lápityúk |
| Indonesian | Kareo padi |
| Japanese | シロハラクイナ |
| Korean | 흰배뜸부기 |
| Malayalam | കുളക്കോഴി |
| Marathi | पांढऱ्या छातीची पाणकोंबडी |
| Norwegian | hvitbrystrikse |
| Odia | ଡାହୁକ |
| Persian | طاووسک سینه سفید |
| Polish | bagiewnik białopierśny |
| Portuguese (Portugal) | Franga-d'água-de-peito-branco |
| Punjabi (India) | ਚਿੱਟ-ਹਿੱਕੀ ਜਲ-ਕੁਕੜੀ |
| Russian | Белогрудый погоныш |
| Serbian | Belolika barska kokica |
| Slovak | kohútec bieloprsý |
| Spanish | Gallineta Pechiblanca |
| Spanish (Spain) | Gallineta pechiblanca |
| Swedish | vattenhöna |
| Telugu | తెల్ల బొర నీటి కోడి |
| Thai | นกกวัก |
| Turkish | Ak Göğüslü Suyelvesi |
| Ukrainian | Багновик білогрудий |
Revision Notes
Shawn M. Billerman revised the Systematics section to align with Clements taxonomy. Chuenchom Hansasuta and Peter Pyle contributed to the Plumages and Molts pages. Brooke K. Keeney standardized the account with Clements taxonomy.
Amaurornis phoenicurus (Pennant, 1769)
Definitions
- AMAURORNIS
- PHOENICURUS
- phoenicurus
The Key to Scientific Names
Legend Overview
Field Identification
28–33 cm; male 203–328 (228.5) g, female 166–225 (196·5) g; wingspan 49 cm. Relatively large, distinctive species; unmistakable, with combination of dark slate gray upperparts and flanks, prominent white face and underparts, and rufous rear flanks, vent and undertail coverts. Female smaller, sometimes with duller bill. Some birds have gray sides of breast strongly barred with olive; partial albinism seems to occur in some areas, e.g., Andamans, Nicobars, and Wallacea. In nonbreeding season, male's bill becomes olive, washed brown on upper mandible. Immature duller than adult: upperparts more olive brown, white of face obscured by brown feather tips; underparts tinged brownish; rufous areas duller; bill darker and duller. Juvenile similar but white of underparts, especially chin, throat and sides of breast, more heavily tinged dull brown; bill dark; legs and feet brownish yellow.
Plumages
Rails (Rallidae) have 10 full-length primaries (numbered distally, from innermost p1 to outermost p10), 12 secondaries (numbered proximally, from outermost s1 to innermost s9 and including 3 tertials, numbered distally, t1 to t3), and 12 rectrices (numbered distally on each side of the tail, from innermost r1 to outermost r6). Most rails are diastataxic ( 1 ) indicating that a secondary has been lost evolutionarily between what we now term s4 and s5. Wings are rounded, tail is short and squared, and bill and legs are strong. Geographic variation in appearance is slight to moderate (see Systematics); the following covers all subspecies and is based on descriptions in Ripley (2) and Taylor (3, 4), along with examination of Macaulay Library images. See Pyle (5) and Marchant and Higgins (6) for information on age and sex determination in rallids, including Amaurornis, and see Molts for molt and plumage terminology. Sexes are alike in all plumages; definitive appearance is assumed following the second prebasic molt. Timing of plumages usually relates to timing of breeding and molts (with worn plumages in breeding periods and fresh plumages in non-breeding periods), which can occur year-round among populations of this species (see Breeding: Phenology).
Natal Down
Hatchlings are covered by dense natal down, which is brownish black, grayish black, or black. The down of the femoral feathering can be browner.
Juvenile Plumage
Juvenile Plumage is duller and can be blacker above than definitive basic plumage, the upperpart feathers often diffusely fringed buff or pale brown. The sides of the head are grayish to whitish to mixed sooty and white. The underparts are duller whitish than in later plumages, often tinged grayish. The sides of breast and flanks are more extensively mottled sooty. and the undertail coverts are buff to pale brownish, less richly colored than in later plumages. Juvenile flight feathers are narrower and more pointed or tapered at the tips than basic feathers. See also Bare Parts for age-related differences in bill, iris, and leg colors.
Formative Plumage
Formative plumage resembles Definitive Basic Plumage but averages duller and browner. The upperparts are more olive-brown, the white parts of face can sometimes be mottled with brown feather tips, and the underparts can be tinged brownish and with rufous areas duller. Formative Plumage may best be identified by retained juvenile lower and outer upperwing coverts, contrasting with darker and fresher, replaced formative inner wing coverts, and by retained juvenile primaries, secondaries, and rectrices that appear narrower. more tapered or pointed at the tips, relatively more worn., and paler brown than basic feathers; rectrices may be mixed with formative central and juvenile outer feathers or some may replace all rectrices during the Preformative Molt (study is needed). See also Bare Parts for age-related differences in bill, iris, and leg colors.
Definitive Basic Plumage
The central crown, nape, upperparts, and upperwing coverts are dark slate gray, with the lower back, rump, and uppertail coverts (sometimes the central crown) variably washed olive-brown to brown. Rectrices are slaty olive to dark brown. The forehead, sides of the face, chin, throat, breast, and central abdomen are white, contrasting distinctly with the slaty upperparts from above the eye to the sides of the breast, and less distinctly from the breast to the lower underparts, the sides and flanks sometimes mottled sooty. The rear flanks, femoral feathers, vent and undertail coverts are bright chestnut to rufous. Definitive Basic Plumage averages brighter than Formative Plumage and shows uniformly slate upperwing coverts, and outer primaries and rectrices are broader, more squared, duskier, and relatively fresh, the rectrices uniform in wear without molt limits. See also Bare Parts for age-related differences in bill, iris, and leg colors.
Molts
Molt and plumage terminology follows Humphrey and Parkes (7) as modified by Howell et al. (8). Under this nomenclature, terminology is based on evolution of molts along ancestral lineages of birds from ecdysis (molts) of reptiles, rather than on molts relative to breeding season, location, or time of the year, the latter generally referred to as “life-cycle” molt terminology (9; see 10). In some birds the Humphrey-Parkes (H-P) and life-cycle nomenclatures correspond to some extent but terms are not synonyms due to the differing bases of definition. Prebasic molts often correspond to “post-breeding“ or “post-nuptial“ molts, preformative molts often correspond to “post-juvenile“ molts, and prealternate molts often correspond with “pre-breeding“ molts of life-cycle terminology. The terms prejuvenile molt and juvenile plumage are preserved under H-P terminology (considered synonyms of first prebasic molt and first basic plumage, respectively) and these terms do correspond with those of life-cycle terminology.White-breasted Waterhenexhibits a Complex Basic Strategy (see 8), including complete prebasic molts and a partial-to-incomplete preformative molt, but no prealternate molts, as in other most or all other rails (11, 5), including other Amaurornis rallids (6).
Prejuvenile Molt
Complete, in or near the natal territory. No detailed information on this molt in White-breasted Waterhen.
Preformative Molt
Based on examination of Macaulay Library images, the Preformative Molt can be partial to incomplete, and includes most or all body feathers, a variable number of proximal secondary coverts (but usually not the outer greater coverts), and 1–4 central rectrices in some birds (perhaps occasionally to all rectrices), but no primary coverts, primaries, or secondaries including tertials (11, 6, 5). Study is needed on variation in the extent of this molt in White-breasted Waterhen. In other rallids, Preformative Molts typically occur from 1-3 months following fledging, and can commence on breeding grounds and complete on winter grounds.
Definitive Prebasic Molt
This molt is complete as in most or all other rails, and likely occurs on or near the breeding grounds within 1-2 months following breeding. Molt of remiges is synchronous, resulting in a flightless period during which birds are relatively inactive and are hidden in dense vegetation; there are few to no birds molting primaries among Macaulay Library images of this species. Although replacement of primaries is rapid it may occur proximally (from p10 to p1) in rails (12), opposite to that of most birds but typical of the Gruiformes (13).
Bare Parts
The following is based on published descriptions (2,3, 4) along with examination of Macaulay Library images. See also images under Plumages.
Bill and Shield
The bill changes color by both age and season. In nestlings it is blackish (see images under Natal Down), gradually and variably becoming grayish at the base of the culmen and dusky yellowish to greenish on the lower mandible in juveniles and early during the first year. By the end of the first year (Formative Plumage) the bill can become greenish or greenish with a dusky culmen. In adults it is usually brightish green-yellow with a duskier culmen during non-breeding periods and can become uniromly brighter yellow during pre-breeding and breeding periods. At the base of the culmen a brownish to reddish shield usually develops during the first year, which is smaller and duller (often maroon) than in adults. In breeding adults this shield enlargens to extend up the forehead and becomes bright red, with the red color extending to the base of the culmen and sometimes the base of the lower mandible. During molting and non-breeding periods the shield can reduce in size and become browner. Females may average duller bills and shields, especially during the breeding season.
Iris and Facial Skin
Iris color transitions from blackish in nestlings to brown in juveniles and during the first year, and then brownish red to deep reddish in adults. The bare skin above the bill and around the eye transitions color from gray in nestling and juveniles to red in adults.
Tarsi and Toes
Legs and feet are blackish in nestlings (see images under Natal Down). As juveniles and during the first year they gradually change to grayish or pinkish then dusky yellow and dull yellow. In adults they are dull yellow during non-breeding periods, becoming brighter and often tinged orangish during prebreeding and breeding periods.
Measurements
Linear Measurements
Length
280-330 mm.
Wingspan
490 mm.
Mass
166-225 (196.5) g in females, and 203-328 (228.5) g in males.
Systematics History
Considerable individual variation in size and plumage renders determination of subspecies difficult, leading to various treatments. Here, possible subspecies chinensis (much of mainland range from India east to eastern China) and javanica (Malay Peninsula, Greater Sundas) included within nominate owing to overlap in measurements, but may well be valid; proposed subspecies maldiva (Maldive Islands) also subsumed within nominate, and variabilis (Sulawesi) in leucomelana.
Subspecies
Birds from Sangihe and Talaud Islands (northeast of Sulawesi) normally included in nominate subspecies, but may be closer to other Sulawesi forms; further study needed (14).
Amaurornis phoenicurus phoenicurus Scientific name definitions

Distribution
Pakistan, India, Maldives, and Sri Lanka to east China, Taiwan, Ryukyu Islands, Bonin Islands, and Japan, and south through southeastern Asia and Philippines to Greater Sundas, Christmas Island, and Cocos (Keeling) Islands; northern populations winter to south, ranging west to Arabia. Currently expanding north into Japan and Korea, and east into Micronesia (Cocos, Yap) (15).
Identification Summary
The gray on the head does not extend below the eyes in the nominate .
Amaurornis phoenicurus phoenicurus (Pennant, 1769)
Definitions
- AMAURORNIS
- PHOENICURUS
- phoenicurus
The Key to Scientific Names
Legend Overview
Amaurornis phoenicurus insularis Scientific name definitions

Distribution
Andaman Islands and Nicobar Islands (except central Nicobars).
Identification Summary
Gray on the head reaches only half way over top of head, and the flanks are blackish gray.
Amaurornis phoenicurus insularis Sharpe, 1894
Definitions
- AMAURORNIS
- PHOENICURUS
- phoenicurus
- insularis
The Key to Scientific Names
Legend Overview
Amaurornis phoenicurus leucocephala Scientific name definitions
Systematics History
Described as A. p. leucocephala Abdulali, 1964. Long unrecognized, until Praveen and Khot (16) examined new material, resulting in its recognition.
Distribution
Car Nicobar Island (16).
Amaurornis phoenicurus leucocephala Abdulali, 1964
Definitions
- AMAURORNIS
- PHOENICURUS
- phoenicurus
- leucocephala / leucocephalos / leucocephalus
The Key to Scientific Names
Legend Overview
Amaurornis phoenicurus midnicobarica Scientific name definitions
Distribution
Central Nicobar Islands.
Identification Summary
The gray on the head reaches only half way over top of head, and the underparts are more olive in midnicobaricus.
Amaurornis phoenicurus midnicobarica Abdulali, 1979
Definitions
- AMAURORNIS
- PHOENICURUS
- phoenicurus
- midnicobarica
The Key to Scientific Names
Legend Overview
Amaurornis phoenicurus leucomelana Scientific name definitions

Distribution
Sulawesi, western Maluku Islands (Moluccas), and Nusa Tenggara Islands (Lesser Sundas).
Identification Summary
In subspecies <em>leucomelanus</em> , the gray on head usually extends at least to forehead, and in birds from Nusa Tenggara Islands, to ear-coverts and lores. The extent and shade of gray on flanks is most extensive in <em>leucomelanus</em> , relative to other subspecies.
Amaurornis phoenicurus leucomelana (Müller, 1842)
Definitions
- AMAURORNIS
- PHOENICURUS
- phoenicurus
- leucomelan / leucomelana
- Leucomelana
The Key to Scientific Names
Legend Overview
Distribution
Pakistan, India, Maldives, and Sri Lanka to east China, Taiwan, Ryukyu Islands, Bonin Islands, Japan, south through southeastern Asia and Philippines to Greater Sundas, Christmas Island, Cocos (Keeling) Islands, Andaman Islands, Nicobar Islands, Sulawesi, western Maluku Islands (Moluccas), and Nusa Tenggara Islands (Lesser Sundas).
Northern populations winter to south, ranging west to Arabia. Currently expanding north into Japan and Korea, and east into Micronesia (Cocos, Yap),
Habitat
Reedy or grassy swamps, marshes, tall grass with reeds and shrubs, bamboo stands, wet scrub, rice fields and sugar cane, sewage ponds, and shores of rivers, ponds, ditches and lakes; also thick forest (Andaman and Nicobar islands), forest edges and clearings, mangrove swamps, and scrub and bushes far from water. Often occurs close to human habitation, e.g., at village ponds, and enters compounds and public parks; runs about under roadside hedges. Occurs from lowlands up to 1500 m, to 2000 m in Nilgiri Hills, southwestern India.
Movement
Resident throughout most of range, but northern populations winter south to Nusa Tenggara Islands (Lesser Sundas), occurring west to Arabia, where 13 recorded in Oman (October–January), 1 United Arab Emirates (November), and 1 Yemen (March). Wintering birds occur in Sumatra November–April. Extent of migration from northernmost breeding areas and seasonality of occurrence not clear; current expansion northwards into Japan indicative of mobility of species; recorded east to Bonin Islands.
Diet and Foraging
Worms, molluscs, insects (beetles, grasshoppers, etc.) and their larvae, spiders and small fish; also grass seeds, and shoots and roots of marsh plants. Usually not particularly shy, and often seen out of cover. Gleans from ground ; pecks seeds from standing grass. Forages in the open along water margins; feeds on beaches at low tide (Maldives).
Sounds and Vocal Behavior
Characteristic very loud call comprises variety of roars, grunts, croaks and chuckles, followed by a monotonous “kru-ak, kru-ak, kru-ak-a-wak-wak”; calling may continue for 15 minutes or more; calls most in early morning and in evening, also through night; often calls from perch in top of shrub or bamboo. Very vocal during breeding season; silent thereafter.
Locomotion
Climbs in bushes and trees, swims, and occasionally dives.
Breeding
India and Pakistan, June–October (southwestern monsoon); Andaman and Nicobar islands, June–July; Ryukyu, April–October; Sumatra, January, May, July, September, November; Java, all months except April, October, and November; Nusa Tenggara Islands (Lesser Sundas), downy chicks April–May; in Borneo, and probably on other tropical islands, may breed in almost any month. Normally seen singly or in pairs, sometimes in groups of up to five; presumed monogamous, with pair-bond at least for duration of breeding season when apparently territorial. Nest a shallow cup-shaped pad of twigs, creeper stems and Typha leaves, placed on ground in grass or tangled undergrowth at margin of pond, ditch or flooded rice field, or concealed in shrub, bamboo clump or Pandanus palm thicket up to 2 m above ground, sometimes far from water. Eggs 4–9; incubation 20 days, by both sexes; downy chick black , with blackish bill and legs ; chicks leave nest soon after hatching; fed and cared for by both parents. Probably multi-brooded in Ryukyu Isalnds, which has aided rapid expansion there in recent years.
Conservation Status
Not globally threatened (Least Concern). Formerly regarded as common throughout range. Currently common in Borneo, Sumatra, Java, and Bali, and fairly common though local in Philippines. Accidental in Japan before 1970, after which has undergone major range extension, becoming common breeding resident in Ryukyu Islands (Okinawa and in Yaeyama islands); now so common on Iriomote-jima that is thought to be ousting Slaty-legged Crake (Rallina eurizonoides); expansion northwards is continuing, and now also occurs fairly regularly on main Japanese islands, having bred on Kyushu. It has also expanded into Micronesia where apparently arrived to Woleai Atoll, Yap, in early 1970s and subsequently colonized neighboring atolls; recorded also from Palau, in 2009 (15).
About the Author(s)
Chuenchom Hansasuta received a Doctor of Dental Surgery from Chulalongkorn University. During her long and distinguished career in dentistry, she studied and practiced in places such as Thammasat University (Thailand), State University of New York at Buffalo (USA), and University of Connecticut Health Center (USA) and retired in 2020. Chuenchom always had an intense curiosity for birds that over time, evolved into an acute interest in plumages. She has long been active in education and volunteering, becoming chairwoman of The Flyway Foundation and actively engaging and educating the public in the study of birds and their plumages.
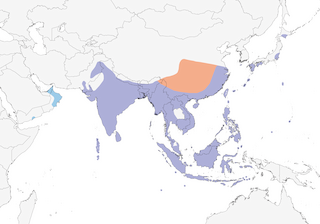
- Year-round
- Migration
- Breeding
- Non-Breeding

















