








Yellow-faced Honeyeater Caligavis chrysops Scientific name definitions
Text last updated January 21, 2013
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Catalan | menjamel caragroc |
| Dutch | Geelmaskerhoningeter |
| English | Yellow-faced Honeyeater |
| English (United States) | Yellow-faced Honeyeater |
| French | Méliphage à joues d'or |
| French (France) | Méliphage à joues d'or |
| German | Dreistreifen-Honigfresser |
| Japanese | キホオコバシミツスイ |
| Norwegian | gulstripehonningeter |
| Polish | maskowik pręgolicy |
| Russian | Желтолицый медосос |
| Serbian | Žutoliki medojed |
| Slovak | medárik topásový |
| Spanish | Mielero Carigualdo |
| Spanish (Spain) | Mielero carigualdo |
| Swedish | gulkindad honungsfågel |
| Turkish | Sarı Yanaklı Balkuşu |
| Ukrainian | Медник жовтощокий |
Caligavis chrysops (Latham, 1801)
Definitions
- CALIGAVIS
- chrysops
The Key to Scientific Names
Legend Overview
Originally Appeared in

Field Identification
15–17·5 cm; male 12–20·5g and female 15–20 g (nominate), two males 17–17·5 g and female 15–16 g (samueli), male 14–17·2 g and female 13·8–15·8 g (barroni). Nominate race is greyish-brown to olive-brown above, with diffuse darker mottling or streaking on top of head, hindneck, mantle, back and scapulars; broad curving yellow stripe from gape backwards below eye and across upper ear-coverts to meet small white patch on rear ear-coverts, and bordered above by black eyestripe (broken above rear edge of eye by small yellow to off-white patch) and below by black submoustachial stripe; uppertail brown; upperwing-coverts greyish-brown to olive-brown, broad off-white tips on outer median and secondary coverts and olive tips on primary coverts; remiges brown with fine olive outer edges (forming faint olive panel); chin, throat and underbody pale greyish-brown, slightly darker on breast, with slightly darker streaking on chin and throat and paler off-white streaking elsewhere; undertail greyish-brown, underwing pale rufous with darker, brown, trailing edge and tip; iris dusky blue, brown in some (possibly immatures); bill black; legs dark grey. Sexes alike in plumage, male slightly larger than female. Juvenile is very similar to adult, but paler and unstreaked above, rump and uppertail-coverts browner, underparts paler (breast unstreaked), bill black with orange-brown tip, gape may be slightly more yellow at least initially. Races vary little: samueli is significantly smaller than nominate, slightly paler and less mottled above, with more off-white belly; barroni is very similar to previous, but slightly paler above and below.
Systematics History
Editor's Note: This article requires further editing work to merge existing content into the appropriate Subspecies sections. Please bear with us while this update takes place.
Sister to the pair C. obscura and C. subfrenata (1). Validity of poorly differentiated race barroni requires confirmation. Three subspecies provisionally recognized.Subspecies
Caligavis chrysops barroni Scientific name definitions
Distribution
Caligavis chrysops barroni (Mathews, 1912)
Definitions
- CALIGAVIS
- chrysops
- barroni
The Key to Scientific Names
Legend Overview
Caligavis chrysops chrysops Scientific name definitions
Distribution
Caligavis chrysops chrysops (Latham, 1801)
Definitions
- CALIGAVIS
- chrysops
The Key to Scientific Names
Legend Overview
Caligavis chrysops samueli Scientific name definitions
Distribution
Caligavis chrysops samueli (Mathews, 1912)
Definitions
- CALIGAVIS
- chrysops
- samueli
The Key to Scientific Names
Legend Overview
Distribution
Editor's Note: Additional distribution information for this taxon can be found in the 'Subspecies' article above. In the future we will develop a range-wide distribution article.
Habitat
Dry, open sclerophyll forests and woodlands dominated by Eucalyptus, including savanna and riverine woodlands, with understorey varying from dense to sparse shrubs, including heath, to sparse open grasses; less often in forests and woodlands dominated by Angophora, Acacia, Banksia, casuarina or cypress-pine (Callitris) and, in NE Queensland (race barroni), swamp-woodland dominated by Lophostemon. Sometimes in wet sclerophyll forests, more so when these heavily flowering. Tends to prefer mature forests and woodlands. Commonly also in coastal heathland or shrubland, particularly with flowering Banksia; and often in urban and suburban parks and gardens, and even found along streets. Also in remnant patches of forest or woodland in agricultural landscapes. Sometimes in mangrove associations, and in plantations of exotic pines (Pinus) and orchards. Occasionally in rainforests (but apparently not in NE Queensland). From coasts to high-altitude subalpine zone.
Movement
Nominate race partly migratory; isolated breeding populations of NE Queensland (barroni) and South Australia (samueli) appear sedentary. Part of nominate race migrates to and from SE Australia, N passage in autumn (Mar–May) and return in late winter-spring (S passage less conspicuous). Main stream follows Great Divide and E coast, on N passage tending NE during early autumn, then NW from about SE Queensland during late autumn-early winter; leaves Great Divide of N Victoria-S New South Wales and its tablelands in winter and moves to N New South Wales and inland, with decrease noted in coastal Victoria, and estimates of at least 20% of population moving, most between Mar and Jun. Migrates by day, in small to large flocks moving in one general direction (though can be seen to move in opposite direction to expected passage at some sites), usually at treetop level; migratory flocks usually of fewer than 100 individuals but can be larger, and several records of flocks of c. 1000. Large numbers observed on N passage, typically in successive flocks, e.g. at Wellington Point (SE Queensland) mean of 555 birds/hour (26–2355). Sometimes travels in mixed flocks, often with Melithreptus lunatus. Vagrant on islands of Bass Strait (mostly King I) and once on mainland Tasmania; vagrant at Iron Range, in far NE Queensland. Doubtful report of two individuals in New Zealand.
Diet and Foraging
Wide range of items, including nectar (often of Eucalyptus, Banksia) and pollen, fruit and seeds, manna, and lerp and honeydew; also arthropods, mainly insects, also some spiders (Araneae) and millipedes (Diplopoda). Primarily arboreal, foraging at all levels, also aerially and, occasionally, on ground; proportion of time spent at different levels varies among studies, but often in subcanopy and canopy. Forages among foliage and at flowers of trees, shrubs and mistletoe (Loranthaceae); less often on bark of trees. Gleans from foliage and bark; probes various substrates, including flowers (for nectar) and bark (for invertebrates, such as insect and their products); sallies to capture prey in air or from foliage. Proportions of different methods vary, but gleaning usually constitutes a large proportion. Active, conspicuous. Tends to forage singly, in twos or in small loose groups of up to ten individuals; larger flocks during migration. Occasionally in mixed flocks dominated by insectivorous birds.
Sounds and Vocal Behavior
Typical daytime song 1·8–2·6 seconds in duration, of 4–6 brief syllables, e.g. “chick-up”, at maximum of 15–17 per minute; infrequent throughout day, slightly more often in first hour of daylight and towards sunset. Dawn song a social chorus starting 35–20 minutes before sunrise and stopping 14–8 minutes before it, a major part of dawn chorus Aug–Jan; individual dawn song rendered as “chee-chitty di-chitty-tee-tee”, duration just over 1 second, at rate of 12–16 per minute. Countersinging, possibly between males, frequent. Calls include repeated, somewhat plaintive but harsh “chip” or “cheep”, uttered on autumn migration, in flight and while perched, migratory flocks producing distinctive chattering; harsh “kheer”. Snaps bill in aggression.
Breeding
Season broadly Jul–Mar, with clutches early Aug to late Feb and nestlings reported until early Mar. Nest built by female, often accompanied by male, a fine and fragile, neat woven cup, sometimes with swollen sides or narrower at rim (contents often visible through base or wall), often made of, or covered with, green materials, especially grass or moss, less often including bark or bark fibre, lichens, rootlets, plant stems, fibre or down, leaves, and wool, fur or hair, bound together and to nest plant with spider web, usually lined with very fine rootlets, grass or feathers or other soft plant material, external diameter 3·8–9·5 cm, depth 4·8–10·7 cm, internal diameter 4·3–7·3 cm, depth 3·5–9·5 cm; placed 0·15–25 m (mean 2·3 m) above ground, usually suspended from thin horizontal or vertical fork, or from foliage or shoots, usually among foliage and twigs of live understorey shrub, sometimes in tree or sapling or, less often, in mistletoe, ferns or mangrove, sometimes in dead vegetation, and often near or overhanging water; within a season, second and third nests tend to be higher than earlier ones. Clutch 2–3 eggs, rarely 1 (mean 2·35); incubation by female, period 13–15 days, though said to be as long as 17 days; chicks brooded probably by female alone, fed by both parents, nestling period 11–14 days, once 17 days; fledglings tended by both parents. Nests parasitized by Shining (Chalcites lucidus) and Horsfield’s Bronze-cuckoos (Chalcites basalis), and Fan-tailed (Cacomantis flabelliformis), Brush (Cacomantis variolosus) and Pallid Cuckoos (Heteroscenes pallidus). Success rate for 508 eggs in 235 nests was 0·35 fledged young per nest, and of 401 nests where outcome known 41·9% produced at least one fledged young; in one study, 89% of failures attributed to predation.
Conservation Status
Not globally threatened. Common in most of range. No estimates of total population; recorded densities of 0·01–2·2 birds/ha and 0·25–1·52 pairs/ha. Formerly considered a pest in commercial fruit orchards.
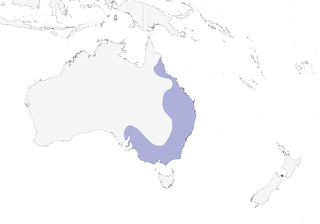
- Year-round
- Migration
- Breeding
- Non-Breeding

eBird and Macaulay Library

eBird Status and Trends

















