











Whooper Swan Cygnus cygnus Scientific name definitions
- LC Least Concern
- Names (48)
- Monotypic
Text last updated August 15, 2016
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Afrikaans | Singswaan |
| Albanian | Mjelma qafëdrejtë |
| Arabic | تم صافر |
| Armenian | Ճչան կարապ |
| Asturian | Cisne glayador |
| Azerbaijani | Harayçı qu |
| Basque | Beltxarga oihularia |
| Bulgarian | Поен лебед |
| Catalan | cigne cantaire |
| Chinese | 黃嘴天鵝 |
| Chinese (Hong Kong SAR China) | 大天鵝 |
| Chinese (SIM) | 大天鹅 |
| Croatian | žutokljuni labud |
| Czech | labuť zpěvná |
| Danish | Sangsvane |
| Dutch | Wilde Zwaan |
| English | Whooper Swan |
| English (United States) | Whooper Swan |
| Faroese | Svanur |
| Finnish | laulujoutsen |
| French | Cygne chanteur |
| French (France) | Cygne chanteur |
| Galician | Cisne bravo |
| German | Singschwan |
| Greek | Αγριόκυκνος |
| Hebrew | ברבור שר |
| Hungarian | Énekes hattyú |
| Icelandic | Álft |
| Italian | Cigno selvatico |
| Japanese | オオハクチョウ |
| Korean | 큰고니 |
| Latvian | Ziemeļu gulbis |
| Lithuanian | Gulbė giesmininkė |
| Mongolian | Гангар хун |
| Norwegian | sangsvane |
| Persian | قوی فریادکش |
| Polish | łabędź krzykliwy |
| Portuguese (Portugal) | Cisne-bravo |
| Romanian | Lebădă de iarnă |
| Russian | Лебедь-кликун |
| Serbian | Veliki labud |
| Slovak | labuť spevavá |
| Slovenian | Labod pevec |
| Spanish | Cisne Cantor |
| Spanish (Spain) | Cisne cantor |
| Swedish | sångsvan |
| Turkish | Ötücü Kuğu |
| Ukrainian | Лебідь-кликун |
Cygnus cygnus (Linnaeus, 1758)
Definitions
- CYGNUS
- cygnus
The Key to Scientific Names
Legend Overview
Originally Appeared in

Field Identification
140–165 cm; male 7200–15,500 g (1), female 5600–13,100 g (2); wingspan 205–243 cm (3). Female averages smaller. Chief risk of confusion is with C. columbianus, especially those birds (apparently of E origin) that can show the angular head and long bill typical of the present species and also seem to have a longer neck and body than is typical of C. columbianus (4). Juvenile shows greyish feathers and has whitish or less usually pinkish, rather than yellowish, base to bill; this latter may be the last indication of the bird’s age in late spring, by which point the feathers are usually entirely white (5).
Systematics History
Subspecies
Hybridization
Hybrid Records and Media Contributed to eBird
-
Mute x Whooper Swan (hybrid) Cygnus olor x cygnus
-
Trumpeter x Whooper Swan (hybrid) Cygnus buccinator x cygnus
Distribution
Iceland and Scandinavia E to NE Siberia; winters in W & C Europe, around Baltic, North, Black, Caspian and Aral Seas, E to coastal China and Japan.
Habitat
Breeds in northern zones, on pools and lakes of shallow fresh waters, preferably in wooded country and only rarely in tundra but this habit is increasing at present (2), up to 700 m in Iceland (2); also on slow-flowing rivers and sheltered coasts. Prefers pools with abundant emergent vegetation, e.g. Arctophila fulva, Equisetum and Carex (2). Non-breeders typically spend summer at lakes, on river channels or coastal bays (2). In NW Europe now winters mainly on low agricultural land , generally not far from coast but always with ready access to areas of open water for roosting; more traditionally and elsewhere, spends winter on freshwater lakes and marshes, brackish lagoons and coastal bays (2).
Movement
Migratory. Part of Icelandic population (typically 500–1300 birds) (2) remains in winter, varying according to weather and food supply (2). Migrates southwards in Sept–Oct (2) to temperate areas, sporadically in more southern latitudes in cold winters; departure for breeding areas commences in Mar–Apr (2), but occasionally remains well to S of breeding range during late spring/early summer, e.g. one early Jun record from N Turkey, where virtually of the comparatively small wintering population have usually departed by late Apr (6). Four main breeding populations—Iceland, N Europe, C Russia and E Russia—winter in different areas, with apparently little interchange, although Icelandic birds have been reported in continental Europe at this season and Finnish breeders in SE England (2): Icelandic population winters mainly in Britain and Ireland (where move readily between England, Scotland and Ireland during course of season), whereas most of those breeding in Finland winter on mainland Europe, especially in Denmark, Norway, Sweden and Schleswig Holstein (Germany), as well as in the Netherlands, the Baltic countries and C Europe, rarely S to N Spain (7), while majority of those wintering in Black Sea and E Mediterranean region probably breed in W Siberia, but those in the area between the Caspian Sea and L Balkash probably originate from C Siberia, and finally those in far E of breeding range winter in Japan , Korea and E China (2). Migration routes in Asia poorly known, but two known in Far East: those from lowlands of NE Russia move through Kamchatka Peninsula and Kuril Is to Japan (on return, from N Honshu, via S Hokkaido, NE Hokkaido and Sakhalin, to the lower Amur R) (8), while those in Kolyma basin suggested to follow W coast of Sea of Okhotsk; also, birds wintering in N Japan (NE Honshu and E Hokkaido) have been satellite-tracked to presumed breeding areas along the middle reaches of the Indigirka R and the lower Kolyma R, in NE Siberia, with stopover sites in Sakkhalin, the lower Amur R and northern coastal areas of the Sea of Okhotsk (9); however, it is thought that the large numbers wintering in Japan must include at least some from as far W as C Siberia (2). Departure from Japan in spring begins in mid–late Mar and takes 26–75 (mean 53·4) days to reach the breeding areas, with the distance travelled being 1600–4000 (mean c. 2935) km (8). Whereas most birds moult close to the breeding grounds, the recently established southernmost breeding populations in Europe appear to perform moult migrations, with at least some of those young bred in Latvia and Estonia moving to Finland and even Arctic Russia to moult, generally leaving before 20 Jun and returning after mid Sept. and moving up to 1455 km (10). In areas where it occurs sympatrically in winter with C. columbianus, the present species tends to arrive earlier but leave later (2). Migratory flights, even over sea, apparently usually at low altitude (11) (maximum confirmed was flock flying at c. 1850 m, but unconfirmed record at 8200 m) and species can cover long distances rapidly, e.g. minimum flight time from Iceland to Scotland just 12·7 hours, with maximum air speeds of c. 97 km/hour (2), whereas during adverse weather conditions, for example strong headwinds, species can take up to four days to make same journey (12), resting on the sea for comparatively long periods (11). Climate change is suspected to be already influencing migratory behaviour in this species, for instance at a stopover site in SC Sweden, the birds are likely to arrive earlier in spring and stay longer, which in turn will probably also result in increased grazing pressure and risk of damage to crops (13), while in the UK data from the period 1972–2008 reveal that earlier departure in late winter/spring is more pronounced for the first 50% of the population than the last 10% of departing individuals, and this pattern is associated with an increase in Feb temperatures on the wintering grounds, which in turn promotes grass growth, enabling the birds’ earlier departure (14). Vagrant to USA (more or less regular in small numbers, < 50 birds, in winter on Aleutians and Pribilofs, with additional records between British Columbia and C California) (15), Taiwan (3), Afghanistan (on border with Iran) (16), Morocco (17), Pakistan and Syria (18).
Diet and Foraging
Almost entirely vegetarian, mainly aquatic plants (Zostera, Ruppia, Potamogeton) and grass during the breeding season, complemented with grain, potatoes (sometimes parasitized from other waterfowl, e.g. Aythya ferina, A. fuligula and Common Coots Fulica atra) (19), acorns and other vegetable matter in winter; also, observed feeding on emerging chironomids in Iceland in spring and freshwater mussels (Mytilus edulis, Unio pictorum) ingested in large numbers in Denmark in winter (2). During breeding season, at least in Finland and Iceland, horsetails (Equisetum), especially E. fluviatile, are important, while sedges (Carex) and common cottongrass (Eriophorum angustifolium) are also commonly selected in these regions (2); also mentioned are Zostera, Ruppia and Potamogeton in Denmark, Chara, Potamogeton and Canadian pondweed (Elodea canadensis) in Scotland, and Ghyceria, Equisetum and marsh yellowcress (Rorippa palustre) in SE England (2). In China, now feeds mainly on wheat, kelp and eelgrass, with wheat accounting for 99·67 % of items in winter and 90·17 % in early spring, and it seems that the principal food item taken formerly, eelgrass, has now been replaced by wheat (20), while in Korea and Japan the species also feeds to some extent on rice (Oryza sativa) fields in winter (21). Practice of feeding on winter crops such as potatoes first noticed in 1940s in NW Europe, where large numbers of swans (up to 60% in S Sweden, where this habit first noted as recently as 1970s) (22) now feed on arable fields, including cereal stubble, winter cereals and oilseed rape, but recent UK surveys found that just 7–13% feed in such sites, albeit in larger flocks, whereas the remainder feed on improved or flooded pastures, or on permanent inland waterbodies (23). Forages on water, by head dipping or upending , or grazing on land . Supplementary feeding , organized both by general public and local government authorities, occurs at most Japanese wintering sites (2).
Sounds and Vocal Behavior
Whooping call , from which species’ English name derives, is bisyllabic, often used repeatedly and deeper and harsher than more musical C. columbianus; strength and pitch of notes vary with social context, being loud and persistent during aggressive encounters and “Triumph Display”, but gentler in contact between pair members and families (2). Also calls prior to take off, becoming louder immediately before flight (2). Cygnets produce high-pitched squeaking notes when distressed, but softer notes in general contact (2).
Breeding
Starts late Apr–May in Iceland and Fennoscandia, but from mid–late May in Russia and even into June at some latitudes, with eggs typically hatching in mid Jun to early Jul, exceptionally late May (2). Mainly long-term monogamous, but 5·8% of pairs involved new partner, while original pair member was still living and divorce (which is much more frequent than in C. columbianus) is apparently unrelated to breeding success (2); pair formation occurs mainly in spring/summer, only occasionally in winter (2). Some 60–70% of birds that return to breeding grounds remain in non-breeding flocks in any given season (2), while among territorial pairs 8–10% do not lay or fail to hatch eggs (24). In solitary pairs in well-defended territories (including against such birds as Anser fabalis) (25), in Iceland typically 1·7–2·5 pairs/10 km2 or 3·9–6·6 pairs/10 km2 in less suitable habitat, and pairs may breed as little as 50 m apart in marshy sites with particularly abundant vegetation (2); nests on large mound of plant matter , mainly moss and lichens. May attempt second clutch if first is lost sufficiently early in nesting season (2). Clutch 4–5 eggs (2–8 (2), exceptionally up to 12) (26), laid at two-day intervals, creamy white, size varies with altitude, at least in Iceland, in lowlands 93–126 mm × 61–79 mm, in highlands 99–121 mm × 64–75 mm, mean mass 328 g (Iceland) or 331 g (Europe) (2); incubation 31–42 (mean c. 35) (2) days by female guarded by male (2); chicks have pale silvery grey down above , white below, hatch asynchronously (2); fledging c. 87 days (c. 80 in captivity) (2) and young remain with adults throughout first winter and spring migration, but do not return to their parents’ breeding territories (2). Sexual maturity at c. 4 years, but two records of females breeding at just one year old and 25% of those in second winter appeared to be paired, whereas some birds do not breed until seven years old (2); oldest wild bird over 22 years (25 years in captivity) (2). Breeding success generally known from studies of winter flock composition, which at three sites in British Isles in 1980s comprised 8·6–22·4 juveniles (2), and is typically higher in summers following mild winters (27). Fewer adults attempt to breed during cold springs, while birds in S Finland laid larger clutches and suffered lower cygnet mortaity than those in N of country (2); in Iceland, females tended to lay more eggs in lowlands, with mean clutch size of 4·7 eggs, versus 3·8 eggs per clutch in highlands, whereas mean brood size (3·1 cygnets) was identical at highland and the lowland sites, but highland cygnets were significantly smaller and lighter (24). Mean annual mortality after fledging c. 17 % in one study, while in Finland it was 14·9%, with rates of 30%, 25% and 12% for first-year, second-year and older birds, respectively, whereas in Iceland minimum survival rate for first-winters was 82·6%, declining to 80·1% for those aged 2–6 years and 78·3% for swans at least eight years old (2); some of difference in these rates implied to fact that Icelandic birds make consistently longer sea crossings (c. 800 km) on migration (2).
Conservation Status
Not globally threatened (Least Concern). Now protected in many range states, including Iceland (since 1885), Japan (1925), Sweden (1927), UK (1954) and Russia (1964) (2). Slight increase in overall numbers since late 1960s/early 1970s. Apparently abundant in parts of Siberia. The four different subpopulations are most easily censused on their wintering grounds. That in Iceland (where illegal hunting appears to be ongoing, with additional problems being collisions with overhead powerlines, adverse weather conditions and predation) numbered 20,856 in Jan 2000, representing an increase on the Jan 1986 census, which registered 16,700 birds, and this subpopulation appears to have definitely increased since 1960s, when c. 4000 were estimated to winter in Britain and 4000–6000 in Ireland (2). Even in the British Isles, some illegal shooting is obvious (28). The NW European subpopulation, which breeds in N Scandinavia and NW Russia has also shown a significant increase since the mid 1960s; it numbered an estimated 25,000 by the mid 1980s and a special survey in 1995 located > 20,000 in Denmark, 14,000 in Germany, 8000 in Sweden (where numbers have increased since the 1970s) (22), > 5000 in Norway and 3000 in Poland, and considered that the entire subpopulation might then number c. 59,000 birds (2). In Finland, where close to extinction in 1950s (2), the species has increased by 11% annually since 1950, with 5000 birds at end of 1980s, while range and numbers are also on increase in neighbouring Sweden, from 20 pairs in 1950 to 310 pairs in early 1970s and 2775 pairs in 1997 (2); breeding range has also spread to Estonia (30–40 pairs), with > 30 pairs in Latvia, 20–40 pairs in Lithuania, 70–80 pairs in Poland (26) (increasing to 120 pairs in 2015) (29) and 20–30 pairs in Germany (2). A pair successfully bred in E France in 2012 (30). Further E numbers become harder to estimate, but 9800 pairs were estimated to breed in C Russia in the early 1980s (where population considered stable) and midwinter counts from the Black Sea and E Mediterranean region placed this subpopulation at c. 17,000 birds (of which c. 3300 winter in Romania), whereas mid 1990s estimates put the W Asia wintering population at c. 20,000 individuals, by which time it was speculated to be declining (2), although numbers in Turkey have been increasing since late 1980s, with the largest-ever winter being 1200 in 1998 (6), while counts in Iran generally range between c. 60 and c. 1500 individuals, mainly in N of country, with an exceptional 26,431 recorded in Jan 2003 (31). Overall, c. 100,000 reckoned to winter throughout W Palearctic, perhaps 60% of global population. Significant concentrations of thousands of wintering birds in Japan (where winter range has shrunk due to wetland drainage since 1920s) (2), South Korea and Turkmenistan; midwinter censuses in Japan, China and Korea suggested that E Siberian subpopulation might have numbered c. 30,000 in mid 1990s, but since then new data indicate that the real total might be double that, with up to 15,000 birds recorded in China in winter, 3500 in Korea and mean wintering totals of 31,000 birds in Japan (2). Small breeding populations in NW China threatened by overgrazing, disturbance and other human activity (2). Some reductions or local extinctions, e.g. formerly bred in S Greenland, Faeroes (until 17th century) and on Orkney Is (until 18th century) (2); declines caused by excessive hunting and also habitat destruction, especially in Asian part of breeding range.
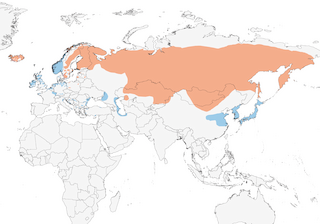
- Year-round
- Migration
- Breeding
- Non-Breeding


















